Vol. 2¯ - XXVII.
Melanociti, melanofori e melanosomi
In seno ai vertebrati esistono almeno due tipi distinti,
ma ontogeneticamente correlati, di cellule produttrici di melanina, diversi luno
dallaltro in modo spiccato circa forma, funzione e risposta agli ormoni
ipofisari:
ñ
melanociti
cutanei ed extracutanei, tipici dei vertebrati omeotermi, quali gli
uccelli e i mammiferi
ñ
melanofori,
tipici dei vertebrati a sangue freddo.
Rimangono ancora dei problemi aperti a proposito di altri
tipi di cellule produttrici di melanina, come le cellule di Kupffer del fegato
e le cellule della ghiandola dellinchiostro dei cefalopodi. Nonostante
siano considerati melanociti, queste cellule non originano dalla cresta
neurale. Cosš, se lontogenesi dalla cresta neurale deve rappresentare il
criterio in base al quale definire una melanogenesi vera, ne consegue che gli
ultimi due tipi cellulari occupano una posizione equivoca.
1. Origine embriologica delle cellule melaniche
Sia i melanofori che i
melanociti dei vertebrati derivano embriologicamente da una popolazione
cellulare di melanoblasti
che nascono dalla cresta neurale, da dove migrano per popolare i
vari distretti dellorganismo. I melanoblasti, se raggiungono la loro giusta
sede, vanno incontro a maturazione sotto forma di melanociti e melanofori
funzionalmente attivi. Solo quei tessuti contenenti la cresta neurale, o
cellule da essa derivate per migrazione, possono produrre melanociti deputati
alla sintesi di pigmento.
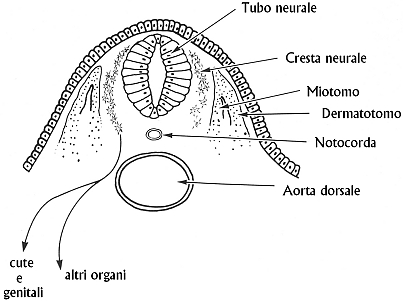
Fig. XXVII. 1.
Migrazione dei melanoblasti
dalla cresta
neurale in un embrione di vertebrato
1.1. Origine dei melanociti
Fu Mary Rawles (1947)
a
scoprire che nel topo, e verosimilmente in tutti gli altri mammiferi, i
melanociti dellepidermide prendono origine dalla cresta neurale, dove le cellule pigmentarie
primordiali derivano da una linea cellulare comune a quella delle cellule che
costituiranno la parte sensitiva afferente dei gangli spinali nonchÕ dei
gangli orto e parasimpatici del neurovegetativo, linea cellulare che Ò pure
condivisa dagli elementi che diventeranno le cellule
C della tiroide
[1]
,
le cellule della midollare del surrene e quelle del glomo carotideo
[2]
.
Una volta giunti nella sede definitiva, i melanoblasti diventano dopa positivi
e sono pronti a iniziare la sintesi melanica.
Attualmente il metodo migliore per identificare le cellule derivate dalla cresta
neurale non consiste nelle metodiche di colorazione, bensš nellimpiego
di anticorpi
monoclonali specifici. Embriologicamente i melanociti riconoscono una
duplice origine:
ñ
lo strato esterno della coppa ottica genera lepitelio pigmentato della retina e del
corpo ciliare, lo strato pigmentato posteriore delliride (o epitelio
pigmentato delliride), forse anche le cellule pigmentate del pecten
ñ
la cresta neurale dÁ luogo ai melanoblasti che migreranno in tutti gli
altri distretti dellorganismo, inclusa la porzione uveale dellocchio
composta da coroide, gran parte del corpo ciliare e superficie anteriore delliride
(o endotelio delliride). Ovviamente dalla cresta neurale originano anche i
melanociti della substantia nigra
[3]
e del locus caeruleus.
Si Ò sempre pensato che la sorgente dei melanociti fosse
rappresentata da un singolo tipo cellulare, cioÒ da quelle cellule che
nascono dalla cresta neurale e che successivamente prendono posto nei vari
tessuti pigmentati. Le differenze morfologiche e funzionali presenti nei
diversi distretti scaturirebbero da interazioni tra melanociti e ambiente.
Tutto ci· Ò in parte vero, in quanto la coppa ottica si stacca
bilateralmente dal tubo neurale.
Ma, nel 1986, Boissy ha osservato che le colture di
cellule della cresta neurale contengono due tipi distinti di melanociti: un tipo produce un
pigmento uguale a quello delle piume che stanno rigenerando, mentre laltro
contiene melanosomi simili a quelli della coroide. Da ci· si pu· dedurre
che, pur avendo la stessa origine, i melanoblasti della cresta neurale si
differenziano giÁ durante la vita embrionale e che tale differenziazione non
dipende dallambiente, bensš da fattori di origine strettamente genetica.
|
sommario |
avanti |
|
|
|