Lessico
Girolamo Fabrizi
d'Acquapendente

![]()
![]()
Girolamo
Fabrizi d'Acquapendente (Acquapendente![]() , VT, 20 maggio 1537 – Padova, 21
maggio 1619) č stato un anatomista, chirurgo e fisiologo italiano. Č
conosciuto anche come Girolamo Fabrizio o col nome latino Hieronymus
Fabricius.
, VT, 20 maggio 1537 – Padova, 21
maggio 1619) č stato un anatomista, chirurgo e fisiologo italiano. Č
conosciuto anche come Girolamo Fabrizio o col nome latino Hieronymus
Fabricius.
La data di nascita č incerta e a volte č riportato l'anno 1533. Dopo gli
studi primari, nel 1550 si recň a Padova, dove si laureň nel 1559. Divenuto
allievo e amico di Gabriele Falloppia (Modena 1523 - Padova 1562), nel 1565
gli succedette, dopo averlo supplito, divenendo professore di anatomia e
chirurgia. Nel 1584 fece costruire, a sue spese, all'interno del Palazzo del
Bo il celebre teatro anatomico![]() che egli stesso inaugurň il 23 gennaio 1594.
che egli stesso inaugurň il 23 gennaio 1594.
Grande didatta, ebbe numerosi allievi e fu maestro, tra gli altri, di Giulio
Cesare Casseri, William Harvey![]() , Adriaan Van Den Spieghel e Werner Rolfinck.
Esercitň anche la medicina pratica, con grande successo e grandi profitti
economici. Ebbe pazienti illustri, dal duca di Urbino al duca di Mantova, da
Carlo de' Medici a Paolo Sarpi, fino a Galileo Galilei di cui fu collega di
insegnamento e amico.
, Adriaan Van Den Spieghel e Werner Rolfinck.
Esercitň anche la medicina pratica, con grande successo e grandi profitti
economici. Ebbe pazienti illustri, dal duca di Urbino al duca di Mantova, da
Carlo de' Medici a Paolo Sarpi, fino a Galileo Galilei di cui fu collega di
insegnamento e amico.
Sotto il profilo scientifico l'Acquapendente diede numerosi contributi
originali e fondamentali in anatomia, embriologia, anatomia comparata (famosa
negli uccelli č la Borsa di Fabrizio![]() ) nonché in chirurgia. Per alcune sue
opere pionieristiche (De formato foetu, De formatione ovi et pulli)
č stato considerato il padre dell'embriologia moderna. In realtŕ l'Acquapendente
fu soprattutto un grandissimo anatomista e fisiologo oltre che un grande
chirurgo.
) nonché in chirurgia. Per alcune sue
opere pionieristiche (De formato foetu, De formatione ovi et pulli)
č stato considerato il padre dell'embriologia moderna. In realtŕ l'Acquapendente
fu soprattutto un grandissimo anatomista e fisiologo oltre che un grande
chirurgo.
Opere principali
Pentateuchus
chirurgicus (1592)
De formato foetu (1600)
De visione voce auditu (1600)
De brutorum loquela (1603)
De venarum ostiolis (1603)
Tractatus anatomicus triplex quorum primus de oculo, visus organo. Secundus de
aure, auditus organo. Tertius de laringe, vocis organo admirandam tradit
historiam, actiones, utilitates magno labore ac studio (1613)
De muscoli artificio: de ossium
articolationibus (1614)
De respiratione et eius
instrumentis, libri duo (1615)
De gula, ventriculo, intestinis tractatus (1618)
De motu locali animalium secundum
totum, nempe de gressu in genere (1618)
De formatione ovi et pulli (1621)
Girolamo
Fabrizi
d'Acquapendente

Monumento di Girolamo ad Acquapendente
Anatomista
e chirurgo nato ad Acquapendente![]() (VT)
attorno al 1533, morto a Padova il 21 maggio 1619. Di nobile famiglia
decaduta, diciassettenne Fabrizi si recň a Padova dove, sotto la protezione
di una famiglia appartenente alla nobiltŕ veneziana, i Lippomano, compě
studi classici, di filosofia e infine di medicina. Fu allievo di Gabriele
Falloppia (Modena 1523 - Padova 1562) di cui seguě i corsi, assistendolo sia
nelle visite ai malati sia nelle dissezioni anatomiche. Attorno al 1559 gli fu
conferito il titolo di dottore in medicina e filosofia.
(VT)
attorno al 1533, morto a Padova il 21 maggio 1619. Di nobile famiglia
decaduta, diciassettenne Fabrizi si recň a Padova dove, sotto la protezione
di una famiglia appartenente alla nobiltŕ veneziana, i Lippomano, compě
studi classici, di filosofia e infine di medicina. Fu allievo di Gabriele
Falloppia (Modena 1523 - Padova 1562) di cui seguě i corsi, assistendolo sia
nelle visite ai malati sia nelle dissezioni anatomiche. Attorno al 1559 gli fu
conferito il titolo di dottore in medicina e filosofia.
Dopo la morte del Falloppia, la cattedra di anatomia restň vacante fino al 1565, anno in cui Fabrizi, che nel frattempo aveva impartito lezioni private, fu incaricato della cattedra di chirurgia con l'implicito accordo di insegnare anche l'anatomia. Il suo stipendio iniziale di cento fiorini veniva aggiornato a ogni riconferma della posizione accademica e, nel 1594, alla quinta riconferma, esso ammontava a millecento fiorini. Fabrizi ebbe successo anche sotto il profilo del prestigio accademico: nel 1584 la sua posizione venne rafforzata dal fatto che il suo insegnamento fu riconosciuto fondamentale. Egli venne quindi eletto membro del Collegio di filosofia e medicina, fino ad allora precluso ai professori di chirurgia, il cui corso era considerato di scarsa importanza e prestigio.
Nel 1600
Fabrizi fu nominato professore supraordinarius di anatomia, con uno
stipendio vitalizio di mille scudi, e nel 1603 professore supraordinarius
di chirurgia. Tutte queste onorificenze non hanno un interesse esclusivamente
biografico, ma rappresentano l'esempio tangibile dell'importanza sempre
crescente che l'anatomia veniva assumendo nell'ambito delle scienze mediche.
Fabrizi insegnň anatomia a Padova per circa quarant'anni ed era ritenuto il
piů grande anatomista del suo tempo. Egli ottenne che l'anatomia
riacquistasse quella dignitŕ istituzionale che giŕ di fatto le era stata
riconosciuta a Padova grazie al giovane umanista e anatomista Andrea Vesalio![]() (Bruxelles
1514 - Zante 1564), che aveva contribuito a trasformare l'anatomia da cruda
pratica qual era, strettamente imparentata alla chirurgia, in parte integrante
e indispensabile della medicina. Inoltre, nel 1594, Fabrizi riuscě a far
costruire un anfiteatro anatomico permanente
(Bruxelles
1514 - Zante 1564), che aveva contribuito a trasformare l'anatomia da cruda
pratica qual era, strettamente imparentata alla chirurgia, in parte integrante
e indispensabile della medicina. Inoltre, nel 1594, Fabrizi riuscě a far
costruire un anfiteatro anatomico permanente![]() , che esiste tuttora, in
sostituzione delle precedenti costruzioni provvisorie.
, che esiste tuttora, in
sostituzione delle precedenti costruzioni provvisorie.
Non si puň dire che Fabrizi sia stato un buon docente, nonostante i suoi indiscutibili successi come ricercatore. Per molti anni, nei rapporti dei rappresentanti degli studenti tedeschi a Padova, vi furono frequenti lamentele per il fatto che Fabrizi non teneva, o non portava a termine, i corsi di anatomia per i quali si era impegnato. La sua didattica era "obscurissima et imperfectissima" e per di piů egli impediva ad altri anatomisti di tenere lezioni private agli studenti. Con il passar degli anni, i rapporti tra Fabrizi e i suoi studenti tedeschi migliorarono a tal punto che nel 1612 il rappresentante tedesco riferiva che Fabrizi era diventato "il nostro comune Maestro".
Il punto forte di Fabrizi stava nella ricerca. Se prendiamo in considerazione la sua lunga carriera, dobbiamo dire che la produttivitŕ scientifica non fu, a dire il vero, eccelsa. Il suo obiettivo era di portare a termine un imponente lavoro, che avrebbe dovuto intitolarsi Totius Animalis Fabricae Theatrum, ma di cui riuscě a concludere soltanto alcune piccole sezioni sotto forma di trattati specialistici, che sperava di vedere un giorno riuniti a formare il Theatrum.
Nel suo
primo trattato, De Visione, Voce, Auditu (1600), Fabrizi espose alcune
linee direttive; lo interessavano in particolare tre aspetti principali
dell'anatomia: la descrizione, il funzionamento e l'utilitŕ delle parti del
corpo. Scrisse che, nonostante Vesalio avesse di gran lunga superato i suoi
predecessori per quanto atteneva l'accuratezza descrittiva, aveva detto ben
poco riguardo all'utilitŕ e al funzionamento delle parti del corpo. Fabrizi
si proponeva di colmare queste lacune. Occorre tuttavia dire che non v'era
nulla di nuovo nel fatto di dare importanza al funzionamento e all'utilitŕ
delle parti del corpo; come Fabrizi stesso ebbe a scrivere, questo approccio
traspariva giŕ in Aristotele![]() e Galeno
e Galeno![]() ed č
chiaro che Fabrizi era orientato piů verso la tradizione che verso
l'innovazione (la suddivisione dell'anatomia in tre parti era stata adottata
anche da molti altri anatomisti del XVI secolo).
ed č
chiaro che Fabrizi era orientato piů verso la tradizione che verso
l'innovazione (la suddivisione dell'anatomia in tre parti era stata adottata
anche da molti altri anatomisti del XVI secolo).
Il risultato di questa sua attenzione particolare al funzionamento e all'utilitŕ era che la pura descrizione di una parte anatomica rappresentava soltanto il primo gradino verso la comprensione della sua utilitŕ. In questo senso Fabrizi, piů degli altri anatomisti del suo tempo, fu portato a integrare l'osservazione con la teoria. Un'altra sua caratteristica saliente era l'interesse per l'anatomia comparata, dove egli seguě chiaramente l'esempio del suo maestro Falloppia. Non di rado Fabrizi č stato considerato l'uomo che ha maggiormente contribuito ad allontanare da Galeno l'anatomia del tardo Rinascimento per indirizzarla verso l'approccio aristotelico. Certamente, il suo ricorrere all'anatomia comparata, l'importanza che attribuiva alla teleologia (tutte le parti del corpo hanno una loro utilitŕ e il fine dell'anatomia dev'essere quello di scoprirla) e la sua predilezione per l'embriologia, rivelano una forte influenza aristotelica che del resto traspare anche dalle sue frequentissime citazioni dell'opera di Aristotele. Fabrizi spesso non sembra tuttavia rendersi conto di questo cambiamento: negli scritti sulle finalitŕ e sui metodi della ricerca anatomica, Galeno viene citato molto spesso, e non in contrapposizione bensě a sostegno di Aristotele.
Uno dei
lavori piů significativi di Fabrizi č il De Venarum Ostiolis (1603)
dove si dŕ una descrizione delle valvole venose. Non fu perň il primo a
scoprirne l'esistenza: nel 1545 Giambattista Canano (Ferrara 1515-1579) riferě
a Vesalio di aver scoperto le valvole dopo aver aperto le vene azygos e
renali, ma non pubblicň mai la sua scoperta. Charles Estienne (1504-1564) e
Jacobus Sylvius![]() (1478-1555) sono altri due anatomisti i cui nomi
sono stati associati alla scoperta delle valvole. Fabrizi fu tuttavia il primo
a occuparsene in dettaglio.
(1478-1555) sono altri due anatomisti i cui nomi
sono stati associati alla scoperta delle valvole. Fabrizi fu tuttavia il primo
a occuparsene in dettaglio.
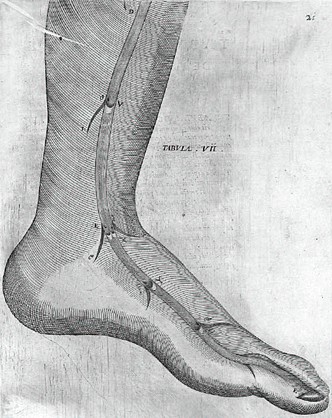
De venarum ostiolis
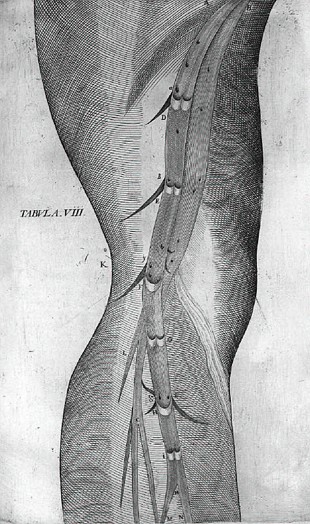
Nel De Venarum Ostiolis, Fabrizi scrisse di aver scoperto per primo le valvole nel 1574: Secondo Salomone Alberti (1540-1600) egli ne fece una dimostrazione ai suoi studenti nel 1579, o nel 1578 secondo quanto afferma Caspar Bauhin (1560-1624). Il ritardo di ben ventinove anni tra la scoperta delle valvole da parte di Fabrizi e la pubblicazione del trattato sull'argomento č da imputarsi a un aspetto tipico del carattere di Fabrizi, che spesso lasciava trascorrere molto tempo prima di pubblicare i suoi trattati in modo da poterli sottoporre a critiche informali e ulteriori aggiornamenti e revisioni. Il ritardo puň anche trovare una giustificazione nel fatto che, durante il XVI secolo, la prioritŕ delle scoperte anatomiche poteva essere attribuita sia per una dissezione effettuata alla presenza di testimoni sia attraverso una pubblicazione.
Le valvole
dei vasi sanguigni avrebbero fornito a William Harvey![]() (1578-1657), il piů famoso degli studenti di Fabrizi, la chiave per poter
giungere alla scoperta della circolazione del sangue. Fabrizi tuttavia collocň
la funzione delle valvole all'interno di uno schema mentale rigido,
tradizionalmente galenico sulla circolazione del sangue. Egli affermň che le
valvole erano state progettate della natura per impedire che le estremitŕ
venissero invase dal sangue. In questo modo, mani e piedi non sarebbero stati
perennemente invasi dal sangue e le restanti parti del corpo potevano avere un
apporto sanguigno pia bilanciato. Una teoria del genere aveva senso soltanto
partendo dal punto di vista di Galeno, secondo cui i diversi distretti
corporei, a seconda dei propri bisogni nutritivi, attiravano il sangue dal
fegato. Le valvole aiutavano le parti centrali e superiori del corpo ad
attirare il sangue impedendo che questo si accumulasse alle estremitŕ. Le
teorie di Fabrizi dovettero essere abbandonate allorquando Harvey, avendo
scoperto che le valvole delle vene superiori erano in realtŕ disposte in modo
da portare il sangue verso it basso in direzione del cuore, giunse alla
conclusione che la funzione delle valvole non poteva essere quella di impedire
un eccessivo irroramento delle estremitŕ. Fabrizi era tuttavia troppo intento
a far conciliare le sue scoperte anatomiche con la teoria tradizionale e ignorň
l'importantissima osservazione di Harvey.
(1578-1657), il piů famoso degli studenti di Fabrizi, la chiave per poter
giungere alla scoperta della circolazione del sangue. Fabrizi tuttavia collocň
la funzione delle valvole all'interno di uno schema mentale rigido,
tradizionalmente galenico sulla circolazione del sangue. Egli affermň che le
valvole erano state progettate della natura per impedire che le estremitŕ
venissero invase dal sangue. In questo modo, mani e piedi non sarebbero stati
perennemente invasi dal sangue e le restanti parti del corpo potevano avere un
apporto sanguigno pia bilanciato. Una teoria del genere aveva senso soltanto
partendo dal punto di vista di Galeno, secondo cui i diversi distretti
corporei, a seconda dei propri bisogni nutritivi, attiravano il sangue dal
fegato. Le valvole aiutavano le parti centrali e superiori del corpo ad
attirare il sangue impedendo che questo si accumulasse alle estremitŕ. Le
teorie di Fabrizi dovettero essere abbandonate allorquando Harvey, avendo
scoperto che le valvole delle vene superiori erano in realtŕ disposte in modo
da portare il sangue verso it basso in direzione del cuore, giunse alla
conclusione che la funzione delle valvole non poteva essere quella di impedire
un eccessivo irroramento delle estremitŕ. Fabrizi era tuttavia troppo intento
a far conciliare le sue scoperte anatomiche con la teoria tradizionale e ignorň
l'importantissima osservazione di Harvey.
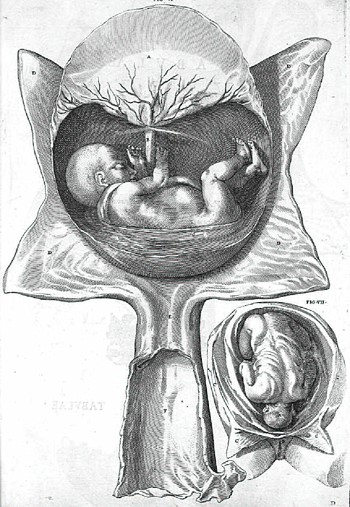
De formato foetu
Anche i trattati di embriologia di Fabrizi influenzarono profondamente Harvey. I due lavori sui quali si basa la fama di anatomista di Fabrizi sono De Formatione Ovi et Pulli (1621) e De Formato Foetu pubblicato nel 1604 ma scritto dopo il precedente. Egli aveva anche scritto, prima dei tre trattati, un De Instrumentis Seminis, che perň non fu mai pubblicato ed č andato perduto. De Formatione Ovi et Pulli č costituito da due parti, la prima delle quali si occupa della formazione dell'uovo, la seconda dello sviluppo del pulcino. Fabrizi iniziň a modificare la classificazione aristotelica, notando che la maggior parte degli uccelli e dei testacei erano ovipari e non venivano quindi, come pensava Aristotele, generati della materia in decomposizione. Fabrizi fu molto prossimo alla conclusione di Harvey secondo cui tutti gli animali derivano da un uovo ("ex ovo omnia") ma riteneva che i mammiferi derivassero dal seme e che alcuni insetti venissero effettivamente al mondo per generazione spontanea.
Fabrizi
passň quindi a considerare l'accrescimento dell'uovo e fu il primo a
comprendere che gli "uteri" del pollo, e cioč l'ovario e
l'ovidotto, svolgono un ruolo cruciale per la formazione e il nutrimento
dell'uovo. Nella seconda parte del trattato Fabrizi discute la strutture, la
funzione e l'utilitŕ dei diversi tipi di uova e successivamente la
formazione, l'accrescimento e il nutrimento del pulcino. Egli riferě di
questi ultimi fenomeni nei termini utilizzati dal metodo scolastico
dell'argomentazione dialettica: adottň l'interpretazione di Aristotele,
secondo cui era la femmina a fornire tutta la materia e il maschio la forma,
per concludere che il seme del gallo č l'agente formativo che si immagazzina
nella borsa cloacale (la "Borsa di Fabrizio"![]() ), mentre
la calaza alla base dell'uovo č la materia con la quale il pulcino si forgia.
Egli ignorň quindi del tutto la cicatricola (o
cicatricula o discoblastula o blastoderma
), mentre
la calaza alla base dell'uovo č la materia con la quale il pulcino si forgia.
Egli ignorň quindi del tutto la cicatricola (o
cicatricula o discoblastula o blastoderma![]() ) e affermň che il tuorlo e l'albume
erano utilizzati per il nutrimento. Il seme veniva quindi tenuto lontano
dall'uovo e Fabrizi ipotizzň che esso fosse in grado di trasmettere il
proprio potere formativo all'uovo attraverso una forza spirituale. Fabrizi
passa quindi a discutere l'ordine secondo il quale le diverse parti del
pulcino si sviluppano. Benché sia in questa occasione molto meno preciso di
quanto non lo fossero Ulisse Aldrovandi (1522-1605) e Volcher Coiter
) e affermň che il tuorlo e l'albume
erano utilizzati per il nutrimento. Il seme veniva quindi tenuto lontano
dall'uovo e Fabrizi ipotizzň che esso fosse in grado di trasmettere il
proprio potere formativo all'uovo attraverso una forza spirituale. Fabrizi
passa quindi a discutere l'ordine secondo il quale le diverse parti del
pulcino si sviluppano. Benché sia in questa occasione molto meno preciso di
quanto non lo fossero Ulisse Aldrovandi (1522-1605) e Volcher Coiter![]() (1534-1576), Fabrizi concluse che doveva esistere una "carena"
("carina") prima ancora che si formassero i primi organi (il cuore
secondo Aristotele, il fegato secondo Galeno).
(1534-1576), Fabrizi concluse che doveva esistere una "carena"
("carina") prima ancora che si formassero i primi organi (il cuore
secondo Aristotele, il fegato secondo Galeno).
Nel De Formato Foetu Fabrizi descrive in che modo la natura ha fornito al feto i mezzi per crescere, per essere nutrito e protetto, per eliminare le sue "scorie" e arrivare con successo alla nascita. Il trattato riguarda i vasi ombelicali, l'uraco, le membrane fetali, le "scorie" del feto, la placenta e l'utero. Fabrizi esordisce fornendo una descrizione precisa di queste strutture, con disegni relativi a una vasta gamma di animali (uomo, cane, gatto, coniglio, topo, ratto, cavia, pecora, bue, capra, alce, capriolo, daino, cavallo, maiale e perfino uccelli e squali). Forně una esauriente descrizione dei vasi ombelicali, del dotto venoso, del foro ovale e del dotto arterioso. Le sue osservazioni erano sempre coerenti con la fisiologia di Galeno secondo cui il feto era nutrito dal sangue proveniente dal fegato della madre (Fabrizi era in netto disaccordo con l'opinione di Giulio Aranzi (1530?-1589) secondo il quale la placenta era l'organo emopoietico del feto).
Il sangue della madre veniva quindi digerito dal feto grazie all'aiuto di forze vitali generatesi nel cuore della madre. Fabrizi considerava il feto come una pianta. nutrita e rifornita di forze vitali della madre attraverso le arterie e le vene ombelicali. Il cuore, il fegato e gli altri organi del feto non avevano alcuna funzione che potesse essere di beneficio al feto stesso, che Fabrizi riteneva completamente dipendente dalla madre. Nelle sue descrizioni del corion e dell'allantoide, e della chiusura del dotto arterioso e del foro ovale dopo la nascita, Fabrizi dimostrň tutta la sua acutezza di osservatore. Egli perň era convinto del fatto che queste sue osservazioni, per essere valide, avrebbero dovuto far comprendere l'utilitŕ delle strutture in questione. Dal momento che questa sua indagine teleologica era basata sui concetti teorici di Aristotele e di Galeno, č del tutto naturale che i suoi risultati fossero, piů che sviluppo di nuove idee, perfezionamento e razionalizzazione di antiche teorie.
Fabrizi
scrisse anche di chirurgia: il Pentateuchus Chirurgicus (1592),
pubblicato senza il suo permesso, e le Operationes Chirurgicae (1619).
Il suo modo di insegnare la chirurgia era conservatore, sia nel contenuto
(egli si avvaleva di fonti antiche e medievali) sia nell'approccio, in quanto
riteneva che un buon chirurgo dovesse tagliare il meno possibile e con la
massima cautela. Fabrizi ebbe ottima reputazione come chirurgo e nel 1607 fu
chiamato a curare Pietro Sarpi alias Paulus Venetus![]() (Venezia
1552-1623) dopo l'attentato alla sua vita; fu ricompensato dei suoi sforzi dal
Senato di Venezia con la nomina a Cavaliere di San Marco.
(Venezia
1552-1623) dopo l'attentato alla sua vita; fu ricompensato dei suoi sforzi dal
Senato di Venezia con la nomina a Cavaliere di San Marco.
Nel frattempo aveva ceduto la cattedra di chirurgia a Giulio Casserio (circa 1552-1616), e nel 1613 abbandonň anche l'insegnamento dell'anatomia per trascorrere il resto della sua vita a occuparsi della pubblicazione dei suoi trattati.
Bibliografia
De
Visione, Voce, Auditu, Padova 1600
De
Locutione et Eius Instrumentis, Venezia 1601
De Venarum Ostiolis, Padova
1603
De
Formato Foetu, Padova
1604
Opera
chirurgica, Venezia
1618
De
Formatione Ovi et Pulli, Padova
1621
Opera
omnia anatomica et physiologica,
Leipzig 1687.
Andrew
Wear
King's College – Aberdeen UK
Fabricius was born in Acquapendente and studied at Padua, receiving an MD in 1559 under the guidance of Gabriel Fallopio where he eventually became professor of anatomy and surgery in Padua from 1562. His pupils during his tenure include William Harvey and Adriaan van den Spieghel.
By dissecting animals, Fabricius investigated the formation of the fetus, the structure of the oesophagus, stomach and bowels, and the peculiarities of the eye, the ear and the larynx. His main claim to fame is the discovery of the membranous folds that he calls "valves" in the interior of veins. He did not know the purpose of these folds; however, they are now understood to prevent blood from flowing backwards. He is also famous for pioneering work that earned him the title of 'The Father of Embryology' in Italian medical science.
Dictionnaire
historique
de la médecine ancienne et moderne
par Nicolas François Joseph Eloy
Mons – 1778




Teatro deriva dal latino theatrum, a sua volta dal greco théatron, da theáomai, io guardo. Il Teatro anatomico di Padova č il piů antico teatro anatomico stabile al mondo, voluto da Girolamo Fabrizi d'Acquapendente nel 1594 e ancora perfettamente conservato. Puň essere visitato al Palazzo del Bo. Il tavolo da dissezione č circondato da sei giri di palchi in noce intagliato. Anticamente, essendo vietata la pratica di dissezione dei cadaveri, i cadaveri venivano issati da un canale sotterraneo attraverso una botola nascosta subito sotto il tavolo.
Il Palazzo del Bo č la sede dell'Universitŕ di Padova dal 1493. L'Universitŕ, fondata da docenti e scolari provenienti da Bologna e pubblicamente riconosciuta a partire dal 1222, con il passare degli anni cominciň a porsi il problema di riunire sotto un'unica sede le diverse scuole aperte nella cittŕ con l'incremento del numero di studenti. Dal 1493 avvenne il graduale trasferimento in un gruppo di edifici che si trovavano in corrispondenza dell'attuale Via VIII Febbraio. Questo gruppo di edifici, in origine residenza della famiglia Papafava, era poi passato sotto la proprietŕ di un macellaio quando questi, avendo fornito delle derrate di carne durante l'assedio della cittŕ, li aveva ricevuti in dono nel 1405 da Francesco I da Carrara, signore di Padova.
Il macellaio vi aveva aperto una locanda (Hospitium Bovis) che aveva come insegna un teschio di bue. La proprietŕ dell' "Albergo del Bove" passň successivamente nel 1539 all'Universitŕ, ma l'appellativo di "Palazzo del Bo" č rimasto fino ai nostri giorni, perché l'ateneo volle conservare nel proprio emblema il teschio di bue.
L'antica locanda fu in un primo momento soltanto riadattata alla nuova funzione, ma nel 1552 fu demolita e furono avviati i lavori di costruzione del nuovo edificio, progettato da Andrea Moroni, che attorno a un cortile di pianta quadrata denominato oggi "cortile antico", sviluppň una doppia loggia a due ordini di colonne sulla quale si aprivano le aule.
Attualmente il Palazzo del Bo č sede del rettorato, della facoltŕ di giurisprudenza e delle aule di rappresentanza delle altre facoltŕ. Le cerimonie ufficiali e la maggior parte delle sedute di laurea si tengono ancora nelle vecchie aule dell'edificio.
Borsa di Fabrizio
o Timo cloacale
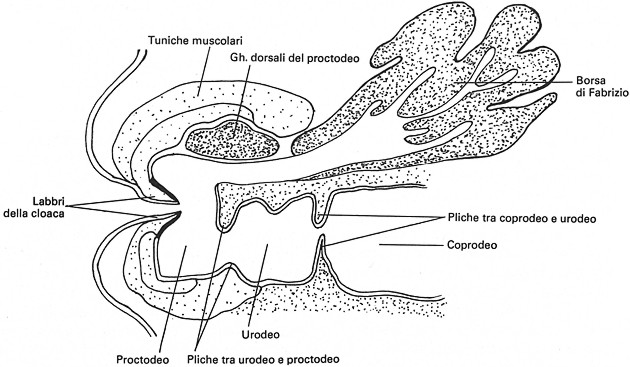
![]()
![]()
La Borsa di Fabrizio o Timo cloacale č un organo tipico degli Uccelli, associato alla parete dorsale della cloaca cui resta connesso mediante un corto dotto che si apre nel proctodeo. Č ben sviluppata nell’animale immaturo (4 mesi in buona parte dei gallinacei); ha forma per lo piů globosa nel pollo, mentre termina a punta nel tacchino e appare come un cieco allungato negli anseriformi.
Dopo alcuni mesi dalla schiusa (5 nei gallinacei, 7 nell’anatra e anche 2 anni nell’oca) va incontro a processi di involuzione, spesso completa all’inizio della maturitŕ sessuale. Molti fattori possono anticipare la regressione dell’organo; tra gli altri vanno ricordati l’ormone adrenocorticotropo, i corticoidi adrenalici, gli steroidi sessuali, alcuni antitiroidei, l’affaticamento, il digiuno e un esaltato esercizio muscolare. Nel pollo la borsa di Fabrizio puň raggiungere il peso di 3 g, anche se esiste una notevole variabilitŕ individuale.
La parete della borsa č composta, procedendo dall’esterno verso l’interno, dalle tuniche avventizia, muscolare e mucosa. Quest’ultima č tappezzata da epitelio cilindrico o pseudostratificato e si solleva in 11-13 pliche principali, ciascuna delle quali appare suddivisa in 6-7 pliche secondarie. La lamina propria della mucosa fornisce una trama per i numerosi follicoli linfatici. Ciascun follicolo presenta tre zone concentriche. La piů esterna (corticale) č fatta di linfociti molto addensati, inframmezzati a cellule reticolari; l’intermedia consta di cellule epiteliali separate dalla corticale dalla membrana basale; l’interna (midollare) presenta cellule reticoloepiteliali e linfociti che sono perň meno compatti. Le cellule reticolari della corticale sono ritenute appartenere al connettivo circostante, mentre quelle della midollare derivano da elementi epiteliali dell’abbozzo embrionale. Nell’asse delle pliche corrono i vasi sanguigni i quali danno poi origine alle reti di capillari che invadono la corticale dei follicoli.
La regressione dell’organo avviene con diminuzione dei linfociti dalla corticale e dalla midollare. Il reticolo epiteliale va incontro a metaplasia dando origine a strutture acinose tappezzate da epitelio cilindrico o pseudostratificato. Segue poi una fibrosi generalizzata con progressiva riduzione di volume dell’intero organo.

De formatione ovi et pulli
Pars Prima – Caput Primum - Podex
Secondo Girolamo Fabrizi, ciň che oggi č un organo linfatico, era invece una borsa in cui finivano il pene e gli spermatozoi del gallo. Si vede che analizzň solamente la cloaca delle galline. Infatti la borsa č presente anche nel gallo, e non solo nel gallo che per motivi contingenti viene montato da altri galli. Gli spermatozoi del gallo trovano accoglienza molto piů in alto, e precisamente 50-80 cm dallo sbocco dell'ovidutto in cloaca: si tratta delle fossette ghiandolari, dove vengono immagazzinati. Le fossette ghiandolari si trovano nel punto di giunzione dell'infundibolo con il magnum.
Sviluppo della borsa di Fabrizio
A 6 giorni di incubazione nell’embrione di pollo si puň apprezzare la formazione di un diverticolo dorsale della parete della cloaca da cui si organizzerŕ la borsa di Fabrizio. Tra il 10° e il 12° giorno compaiono le pliche ancora tappezzate di epitelio indifferenziato. Verso il 12° giorno le cellule epiteliali si proiettano nella lamina propria e qualche giorno dopo danno inizio alla linfocitopoiesi. Secondo alcuni, tuttavia, i linfociti hanno origine all’esterno della borsa nelle pareti del sacco vitellino. Comunque tra le cellule epiteliali compaiono dei linfoblasti basofili che ben presto si differenziano in linfociti grandi, medi e piccoli. Alcune cellule epiteliali si trasformano in elementi stellati con l’apparenza delle cellule reticoloepiteliali. Mentre la linfocitopoiesi č attiva tra le cellule epiteliali e dŕ origine alla midollare dei follicoli, si va formando anche la corticale. Gli elementi mesenchimali della lamina propria proliferano in vicinanza della membrana basale degli abbozzi epiteliali costituendo una rete ove linfoblasti e linfociti appaiono verso ii 15°-16° giorno d’incubazione. Un certo trasferimento di cellule si puň avere dalla midollare alla corticale.
Funzioni della borsa di Fabrizio
Lo sviluppo della borsa di Fabrizio puň essere completamente impedito da una somministrazione precoce di ormoni steroidi sessuali maschili (testosterone, 19-nor-testosterone). Negli animali cosě trattati o in quelli borsectomizzati, le plasmacellule e i centri germinativi dei tessuti linfatici sparsi nell’organismo appaiono deficienti o assenti. Questo fatto dimostra che tali elementi originano per migrazione dalla borsa. Inoltre, l’animale perde piů o meno completamente la capacitŕ a produrre anticorpi e presenta una grave carenza dei meccanismi di immunitŕ umorale, certamente dipendente dalla mancanza delle plasmacellule.
Quali siano i mezzi attraverso cui la borsa di Fabrizio esercita queste funzioni non č ancora del tutto chiaro. Due ipotesi sono oggi accreditate, di queste la prima sembra la piů plausibile.
1) La borsa di Fabrizio č la sorgente di cellule competenti a produrre immunoglobuline; questi elementi si portano precocemente in altri tessuti a cominciare dalla fine del periodo embrionale e nella nuova localizzazione si trasformano in plasmacellule produttrici di anticorpi. Una volta giunte a destinazione tali cellule divengono indipendenti dalla borsa.
2) L’azione della borsa di Fabrizio si eserciterebbe mediante la secrezione di un principio ormonale che, giunto in periferia, determina lo sviluppo della immunocompetenza in elementi linfatici non originati dalla borsa. Questa ipotesi non č stata sinora suffragata da sufficienti prove sperimentali.
Anatomia funzionale degli uccelli domestici
Virgilio Botte & Gaetano Pelagalli
edi.ermes – Milano – 1982
Borsa di Fabrizio
La Borsa di Fabrizio č un organo linfatico primario, scoperto per la prima volta negli uccelli, dove si sviluppano e maturano le cellule B (B da Borsa) che una volta maturati a plasmacellule producono anticorpi. Mentre negli uccelli la Borsa di Fabrizio č situata a contatto e al disopra della cloaca, nei mammiferi nonostante le numerose ricerche non si č riusciti a identificare l'organo linfatico primario che svolga una funzione equivalente. Si ritiene oggi che le cellule B si differenzino dalle cellule staminali presenti nel fegato nel feto e che, dopo la nascita e per tutta la vita, questa funzione venga assunta dal midollo osseo che rappresenta pertanto l'organo borsa-equivalente nel mammifero.
In birds, the bursa of Fabricius (Latin: Bursa cloacalis or Bursa fabricii) is the site of hematopoiesis, a specialized organ that, as first demonstrated by Bruce Glick and later by Max Cooper and Robert Good, is necessary for B cell development in birds. Mammals generally do not have an equivalent organ; the bone marrow is often both the site of hematopoiesis and B cell development. It is named after Hieronymus Fabricius.
Function
The ‘B’ in ‘B cell’ refers to bursa-derived. This is simply because during the 1960s B cells were first defined (and distinguished from thymus-derived T cells) in birds, which have a bursa. A decade later, after examining almost every other organ including the appendix, researchers finally discovered that mammalian B cells develop in the bone marrow and spleen. The fact that ‘bone marrow’, like bursa, starts with a ‘B’ is a coincidence.
The bursa is an epithelial and lymphoid organ that is found only in birds. The bursa develops as a dorsal diverticulum of the proctodael region of the cloaca. The luminal surface of the bursa is plicated with as many as 15 primary and 7 secondary plicae or folds. These plicae have hundreds of bursal follicles containing follicle-associated epithelial cells, lymphocytes, macrophages, and plasma cells. Lymphoid stem cells migrate from the fetal liver to the bursa during ontogeny. In the bursa, these stem cells acquire the characteristics of mature, immunocompetent B cells.
Research history
In 1956, Bruce Glick showed that removal of the bursa in newly hatched chicks severely impaired the ability of the adult birds to produce antibodies. In contrast, removal of the bursa in adult chickens has not much effect on the immune system. This was a serendipitous discovery that came about when a fellow graduate, Timothy S. Chang, who was teaching a course on antibody production obtained chickens from Glick that had been bursectomised (removal of the bursa). When these chickens failed to produce antibody in response to an immunization with Staphylococcus bacteria, the two students realized that the bursa is necessary for antibody production. While some have suggested that this observation was worthy of a Nobel prize, none was awarded, and in fact their initial attempts to publish their findings were thwarted by an editor who commented that "further elucidation of the mechanism ... should be attempted before publication.”
The role of the thymus in the immune response was also identified shortly after the discovery of bursa’s role in antibody responses. In thymectomized animals, the ability to reject allografts, and to mount delayed hypersensitivity responses, was drastically reduced. By the mid-1960s, immunologists were convinced that there were indeed two separate arms of the immune system: one dealing exclusively with the production of circulating antibodies (humoral immunity), and another that is involved in the delayed hypersensitivity-type reactions and graft rejections (cell-mediated immunity).
La bourse de Fabricius (Bursa cloacalis ou Bursa fabricii en latin) est un organe spécialisé dans l'hématopoďčse. Elle est uniquement connu chez les oiseaux. Le nom de cet organe a été donné en l'honneur de Girolamo Fabrizi d'Acquapendente appelé sous le pseudonyme latin de Hieronymus Fabricius.
Il s'agit d'une petite glande (excroissance épithéliale de tissu lymphoďde) présente dans le cloaque des oiseaux, qui semble jouer le męme rôle que le thymus chez les mammifčres, et qui joue un rôle important pour la constitution du systčme immunitaire lors de la phase embryonnaire (maturation des lymphocytes B).Cette glande, découverte par Bruce Glick et étudiée par Max Cooper et Robert Good s'atrophie dans les 6 mois qui suivent la naissance, mais il en reste un vestige fibreux chez l'adulte.
Elle est localisée dans la derničre partie du cloaque des oiseaux, le proctodaeum. Elle est tapissée par la muqueuse intestinale qui forme de nombreux plis. Chaque pli est maintenu par un axe conjonctif constitué de lobules bursiques. Chaque lobule bursique est composé de 2 zones: une zone corticale plus sombre et dense, une zone medullaire moins sombre et dense.
Elle produit des lymphocytes B, B pour Bursa fabricii. En effet cette appellation fut donnée dans les années 1960 lorsque les chercheurs travaillaient sur des oiseaux, pour les différencier des lymphocytes T, dont la maturation nécessite la présence du thymus. Chez les mammifčres les lymphocytes B ne nécessitent que la moelle osseuse pour leur maturation. Selon une étude publiée mi-2007, la taille de cette glande chez les oiseaux pourrait ętre corrélée ŕ leur résistance aux virus de la grippe aviaire faiblement pathogčnes.
L'organo
bursa-equivalente
nell'Uomo e nei Vertebrati
Riapertura
del dibattito e proposta di verifica.
Articolo pubblicato in: EOS. Vol XIX, 1999. n° 1, pag 8-11.
Dopo la scoperta della funzione della Borsa di Fabrizio negli Uccelli (B. Glick 1956), anche nell'Uomo e negli altri Vertebrati si č ricercato affannosamente un organo con tale funzione (Organo Bursa-Equivalente). Sono stati studiati: tonsille, appendice ciecale (specie nel coniglio), placche di Peyer, G.A.L.T. (tessuto linfoide associato al tubo digerente), fegato, midollo osseo. Nessuna di queste formazioni ha caratteri di organo linfoide primario (centrale). Si č arrivati alla conclusione che "nei Mammiferi non esiste un organo bursa-equivalente e la maturazione dei linfociti B avviene, negli stadi piů precoci dello sviluppo, a livello di organi emopoietici quali il fegato fetale, e, successivamente, nel midollo osseo" (vedi note 2, 3). Questa conclusione č in evidente contrasto con l'Anatomia e l'Immunologia Comparate. La Classificazione del Regno Animale (vedi nota 3), sovrapponibile alla Scala della Evoluzione, separa gli Invertebrati dai Vertebrati.
Nei
Vertebrati si passa: dai Pesci agli Anfibi; dagli Anfibi ai Rettili; da
questi: da un lato agli Uccelli, dall'altro ai Mammiferi. Negli Invertebrati
non esistono organi linfoidi, ma solo cellule linfoidi libere. Nei Vertebrati,
dalla Lampreda![]() in su, č sempre presente il Timo: da masserelle irregolari poste dorsalmente
alle fessure branchiali (Pesci) e da masse di tessuto lobulato diffuso
(Anfibi) si passa a due cordoni ghiandolari ai lati del collo (Rettili), a due
strisce di tessuto ghiandolare irregolarmente lobulate lungo tutto il collo
(Uccelli) e infine all'organo pari (Mammiferi).
in su, č sempre presente il Timo: da masserelle irregolari poste dorsalmente
alle fessure branchiali (Pesci) e da masse di tessuto lobulato diffuso
(Anfibi) si passa a due cordoni ghiandolari ai lati del collo (Rettili), a due
strisce di tessuto ghiandolare irregolarmente lobulate lungo tutto il collo
(Uccelli) e infine all'organo pari (Mammiferi).
Seguendo lo sviluppo dell'immunitŕ umorale (vedi nota 4), vediamo che:
- nei Pesci: la Lampreda produce un'immunoglobulina primitiva (formata da una molecola labile a 4 catene);
- nei pesci cartilaginei (Squali) compaiono le plasmacellule e le catene Ig sono legate da ponti disolfuro, ma c'č ancora un'unica classe Ig;
- nei Teleostei si dimostra una primordiale cooperazione tra linfociti T e B, si differenziano gli Antigeni di I e II classe;
- negli Anfibi e nei Rettili compaiono i gangli linfatici e una nuova classe immunoglobulinica, le IgG;
- negli Uccelli compare improvvisamente (?!) la Borsa di Fabrizio e si ha dualitŕ netta delle linee linfocitarie T e B, ma nel Complemento mancano ancora C4 e C2;
- nei Mammiferi compaiono tutte le classi di immunoglobuline: "i Mammiferi sono caratterizzati piů dalla diversitŕ delle classi e sottoclassi di immunoglobuline che da un ulteriore sviluppo dei Linfociti T ". Si avrebbe, nei Mammiferi, un perfezionamento della immunitŕ umorale senza il parallelo perfezionamento dell'Organo che negli Uccelli presiede alla "caratterizzazione" preliminare specifica dei Linfociti B.
Nel Pollo, la Borsa di Fabrizio o Timo Cloacale (vedi note 2, 5, 6, 7) nasce dalla parete posteriore della Cloaca come zaffo epiteliale che si approfonda nel connettivo retrostante, dirigendosi verso il basso. L'intensa proliferazione degli elementi epiteliali porta alla progressiva formazione di un complesso ovoidale, prima pieno, poi contenente numerose piccole cavitŕ irregolari, separate tra loro da setti epiteliali cordoniformi. Nell'ulteriore sviluppo, uno sperone mesenchimale dall'alto si insinua ampiamente tra l'abbozzo della Borsa di Fabrizio e la parete posteriore della Cloaca. In embrioni di 7 giorni, la Borsa di Fabrizio č una formazione autonoma, non comunicante piů con la Cloaca. Da questo momento: la porzione terminale dell'intestino primitivo non subisce piů importanti rimaneggiamenti (rimane la Cloaca definitiva); una depressione ectodermica (il Proctodeo) approfondendosi raggiunge prima la Cloaca (ventralmente) e poi (sul fondo) la Borsa di Fabrizio (embrione di 9 giorni). A sviluppo ultimato, la Borsa di Fabrizio (vedi nota 2, 6, 7, 8, 9) č a forma di sacco, piriforme, raramente cilindrica; č lunga 2 o 3 cm; si trova dorsalmente alla Cloaca, ma comunica col Proctodeo; č fissata dorsalmente mediante un legamento; č rivestita dal peritoneo.
La superficie interna della borsa č a pliche longitudinali piů o meno grosse, che si appiattiscono con l'etŕ. Le sue cavitŕ sono rivestite da epitelio cilindrico, simile a quello intestinale ma privo di cellule mucose. Nella Lamina Propria, subito sotto l'epitelio, si trovano noduli linfoepiteliali in cui si distinguono nettamente un'area centrale chiara (midollare) e un'area periferica scura (corticale). Nel singolo nodulo la midollare č delimitata da una membrana epiteliale continua che, partendo dall'epitelio che tappezza il lume delle cavitŕ, circoscrive un'area grosso modo vescicolare, ovoide, contenente cellule epiteliali lassamente frammiste a grandi linfociti e granulociti. All'esterno della membrana epiteliale, c'č una membrana basale, costituita da una parte centrale (densa e omogenea) e da due strati esterni glicoproteici amorfi. La parte corticale (esterna), costituita da piccoli linfociti e plasmacellule, riveste la zona chiara (midollare) tranne che nella sede d'impianto del nodulo stesso.
La Borsa di Fabrizio per il suo aspetto č nota anche come "Timo cloacale". In un Pollo di 6 settimane č stata descritta "una particolare formazione, costituita da una serie di lamine disposte a cipolla, con predominante struttura linforeticolare. Le singole lamine, separate da uno spazio ricoperto da epitelio cilindrico monostratificato contenente cellule caliciformi, derivano da pieghe epiteliali della parete embrionale della Borsa di Fabrizio e si formano prima della schiusa (8)". La formazione di linfociti nella Borsa di Fabrizio inizia 5 o 6 giorni dopo che nel Timo (vedi nota 6). Da Borse di Fabrizio prelevate dopo la nascita, č stata isolata la bursopoietina (vedi nota 2), un peptide capace d'indurre l'espressione di alcuni antigeni di differenziazione dei linfociti B. La produzione di questo fattore suggerirebbe per la Borsa di Fabrizio una funzione ormonale; ma le ricerche in questa direzione sono state abbandonate perché, utilizzando Borse di Fabrizio embrionarie, non si sono ottenuti risultati positivi. La Borsa di Fabrizio subisce, dopo la nascita, una involuzione direttamente collegata con la maturazione sessuale: comincia ad atrofizzarsi verso il 4° mese di vita (pubertŕ, nei Polli) ed č praticamente scomparsa attorno all'anno (vedi nota 2). La Borsa di Fabrizio č irrorata dall'Arteria Pudenda Communis (vedi nota 5), ramo dell'Arteria Sacrale Media.
Nei Mammiferi l'assetto definitivo della regione caudale č diverso da quello degli Uccelli. Nell'Uomo (vedi nota 9), il setto uro-genitale (o sperone perineale) scende a separare la zona uro-genitale da quella ano-rettale. Nell'embrione umano di 8 settimane lo sperone perineale raggiunge la membrana cloacale, suddividendola in membrana uro-genitale e membrana anale.Tra queste due membrane, il mesenchima dello sperone perineale raggiunge l'ectoderma e, sviluppandosi per formare il perineo, provoca una depressione dell'ectoderma, il Proctodeo. Il riassorbimento della membrana anale, subito dopo, mette in comunicazione il Retto con il Proctodeo (futuro Canale Anale). Dal confronto si vede che l'assetto definitivo della regione anale degli Uccelli corrisponde, fondamentalmente, alla situazione che si ha nell'embrione umano di 4-5 mm, tra la 4a e la 5a settimana di vita fetale. Nell'Uomo, anche nella regione anale, ci si dovrŕ attendere un rimaneggiamento evolutivo in linea con i cambiamenti anatomici che accompagnano il perfezionamento funzionale dei vari organi nei Vertebrati.
Per la comune derivazione dall'epitelio dell'intestino primitivo e tenendo presente la possibilitŕ che la Borsa di Fabrizio abbia una funzione ormonale, vediamo come evolvono Timo e Ipofisi nella Scala Biologica. Del Timo ho giŕ ricordato all'inizio le tappe di organizzazione anatomica, dalla Lampreda ai Mammiferi. Aggiungo che, oltre ad essere organo linfoide primario, il Timo č una ghiandola endocrina; esso deriva da estroflessioni dell'epitelio dell'intestino anteriore primitivo, ma nell'ulteriore sviluppo perde la connessione con la cavitŕ faringea. Dell'Ipofisi ricordo che nella Lampreda, la parte nervosa (neuroipofisi) č costituita da una semplice lamina di tessuto nervoso disposta sul pavimento del Diencefalo (non si forma il prolungamento digitiforme diencefalico che costituirŕ l'infundibulo); la parte endocrina (adenoipofisi) consiste in gruppi di cellule epiteliali sparse attorno al diverticolo ectodermico che si forma per estroflessione dell'epitelio della cavitŕ orale (diverticolo di Ratke). Di particolare importanza č che, nella Lampreda, questo diverticolo rimane sempre in comunicazione con la volta della cavitŕ orale (10).
Dai Pesci piů evoluti, agli Anfibi, ai Rettili, ai Mammiferi, il diverticolo ectodermico perde la connessione con la cavitŕ buccale, diventando 'tasca di Ratke'. Dall'estremitŕ dell'infundibulo e dalla tasca di Ratke proliferano le masse di tessuto che fondendosi danno origine all'ipofisi degli animali piů evoluti (negli Uccelli, manca ancora la pars intermedia). Nell'Uomo l'ipofisi č costituita da "cordoni epiteliali, pieni e cavi, inclusi in logge e concamerazioni connettivali formate da sepimenti della capsula" (vedi nota 11); in essa il sangue venoso arriva dall'Eminenza Mediana attraverso un sistema portale originatosi da "strutture vascolari specializzate, dette "gomitoli" " (vedi nota 12).
L'intera ipofisi umana, nell'adulto, misura mm 10 x mm 13 x mm 6 e pesa da 0,5 a 0,9 grammi (vedi nota 12, 13). Timo e Ipofisi sono solo due esempi: in realtŕ, l'assetto delle principali ghiandole endocrine umane (Ipofisi, Tiroide, Paratiroidi, Timo, Pancreas endocrino) č sempre preceduto, lungo la Scala Biologica, da "stadi" successivi di: cellule sparse; estroflessioni piene o vuote dell'epitelio dell'intestino primitivo; formazioni specializzate comunicanti con la primitiva cavitŕ intestinale; formazioni anatomiche autonome che hanno perduto la connessione originaria con la cavitŕ intestinale. Malgrado i notevoli cambiamenti che si verificano nelle regioni caudali passando dagli Uccelli all'Uomo, ci sono delle strutture che rimangono praticamente immodificate lungo tutta la Scala Biologica: le strutture vascolari (vedi note 3, 7, 14, 15).
In tutti i Vertebrati:
- la Vena Vitellina Sinistra, dopo aver ricevuto anche la Vitellina Dx, circonda la Cloaca (Circolo Venoso Cloacale) e prosegue come Vena Caudale (sempre ben rappresentata dalla Lampreda ai Mammiferi);
- le Aorte Dorsali, dopo la fusione in un tronco unico, mediano, terminano come Arteria Caudale o Sacro-Coccigea.
Nell'Uomo
(vedi nota 16), il ramo terminale dell'Aorta č l'Arteria Sacrale Media (A.S.M.).
Essa nasce dalla parete posteriore dell'Aorta addominale, a livello della 4a
vertebra lombare; scende mediana sulla faccia ventrale di L5, del Sacro e del
Coccige e si esaurisce nella Ghiandola Coccigea di Luschka![]() .
.
La Ghiandola Coccigea, Glomerulo o Paraganglio Coccigeo - G.C., (vedi note 16, 17, 18) si presenta nell'Uomo come un ammasso di 2-3 mm di diametro, globoso o lobulato, impari, mediano, costituito da 3-6 lobuli non sempre bene evidenti. Č costante nel feto, nel bambino e nell'adulto. Si trova immediatamente sotto l'apice del Coccige, un po' spostata ventralmente, compresa tra i fasci trasversali dei muscoli ischio-coccigeo ed elevatori dell'Ano, all'estremitŕ posteriore del Rafe Ano-Coccigeo, in una ganga connettiva molto fitta. In questo connettivo arriva l'A.S.M. che termina al polo superiore della Ghiandola Coccigea: questa "appare come sospesa alla estremitŕ terminale del vaso". La ghiandola č composta da un certo numero di noduli assai irregolari.
Ogni nodulo č composto da un involucro connettivale e da una parte centrale. La parte centrale č composta da un ciuffo di piccoli vasi flessuosi, qua e lŕ rigonfiati, sinusali, attorno ai quali si dispongono ammassi di cellule ghiandolari; in essa si trovano "formazioni cellulari speciali ('corpuscoli lamellari') che per forma e struttura ricordano assai bene i corpuscoli concentrici del Timo". Le cellule che stanno nelle maglie della rete sanguigna (descritte come elementi contrattili vasali che hanno suběto una "curiosa modificazione epiteliale') sono cellule epitelioidi sferiche o poliedriche, a nucleo grosso rotondo od ovoide, con citoplasma perinucleare chiaro, riunite a cordoni e cumuli compatti; contengono granuli di secrezione sicuramente non cromoaffini. La Ghiandola Coccigea "probabilmente č una ghiandola a secrezione interna, di significato assai oscuro (vedi nota 16)"; per il suo colorito rossastro spicca sul tessuto adiposo circostante (vedi nota 17). A differenza dei paragangli addominali (cromoaffini) che derivano sicuramente dall'abbozzo simpatico, " non esiste alcun rapporto genetico fra il simpatico e la Ghiandola Coccigea " (vedi nota 16).
Fin qui abbiamo parlato soprattutto degli Uccelli e dell'Uomo; vediamo cosa avviene andando a ritroso nella Scala Biologica, tenendo presente che i caratteri peculiari della Borsa di Fabrizio sono la derivazione epiteliale dall'intestino posteriore e la ricca e caratteristica vascolarizzazione, derivante dal ramo terminale dell'aorta dorsale. Nei Mammiferi (vedi nota 16), omologhi della Ghiandola Coccigea umana sono i "Glomeruli Caudali", scaglionati uno per vertebra sulla faccia ventrale della coda, lungo l'Arteria Caudale. La loro funzione non č precisata.
Nei Rettili c'č una Cloaca sostanzialmente uguale a quella degli Uccelli (vedi nota 19), e due "ghiandole cloacali", di cui o non si precisa la funzione, o si parla di funzione discussa e di funzione oscura (vedi nota 20). Caratteristica dei Coccodrilli č la Ghiandola paracloacale "grosso organo pari, che sbocca ai lati della fessura cloacale: si tratta di aree ghiandolari affondate a tasca" (vedi nota 21). Proprie dei Cheloni sono delle "insenature dorsali della parete cloacale", sacchi o vesciche anali, che possono essere singole o mancare; la loro funzione č oscura (vedi nota 21). Negli Anfibi la Cloaca ha una dilatazione ventrale bifida che funziona da vescica urinaria; ma il Cieco, corto e ingrossato, puň avere annesso un "cieco dorsale", la cui funzione non č precisata (vedi nota 22).
Nei Pesci:
- in alcuni Teleostei ci sono uno o due "ciechi rettali" (vedi nota 23);
- negli Elasmobranchi, dall'intestino terminale, tappezzato da epitelio pluristratificato, si diparte un cieco rettale o "Ghiandola digitiforme".
La ghiandola digitiforme č un diverticolo a parete sottile che si proietta nel mesenterio dorsale; al suo inizio una valvola impedisce l'entrata degli escrementi; l'epitelio procedendo verso il fondo diventa cilindrico semplice ed č ricco di ghiandole pluricellulari di forma tubulare; č irrorata da due arterie, una anteriore e una posteriore; negli spazi delle ramificazioni ghiandolari c'č una ricca rete di "capillari sanguigni, cui si deve la tinta rosea dell'organo" (vedi note 24, 25), evidente richiamo ai gomitoli caudali ed alla ghiandola coccigea.
Conclusioni
Lo sviluppo progressivo, graduale, senza regressioni, sempre piů affinato dell'immunitŕ umorale nei Vertebrati autorizza a pensare che, nella Scala Biologica, la Borsa di Fabrizio dev'essere stata preceduta e seguita da "equivalenti" piů o meno modificati, da ricercare nella regione sacrale. Confrontando l'assetto definitivo della regione anale negli Uccelli (vedi nota 7) e nell'Uomo, si vede che sono perfettamente sovrapponibili se negli Uccelli si traccia il percorso che lo sperone perineale compie nell'Uomo, andando dal punto di passaggio tra Coprodeo e Urodeo in direzione ventro-caudo-dorsale fino al Proctodeo, tra zona uro-genitale e zona ano-rettale. In questa 'ricostruzione' la Borsa di Fabrizio degli Uccelli occupa ancora il posto che, nell'Uomo, prenderŕ l'ultimo tratto d'intestino posteriore (Retto) per andare a sboccare nel Proctodeo (Canale Anale). Nell'embrione umano, per lo sviluppo del Perineo e per la scomparsa della coda, l'organo bursa-equivalente si sposterŕ lungo il legamento che fissa la Borsa di Fabrizio alle ossa dorsali nel Pollo, finendo come organo anatomicamente autonomo davanti alla punta del Coccige.
Ulteriore conferma dell'equivalenza della Ghiandola Coccigea con la Borsa di Fabrizio viene dalla giŕ ricordata presenza, nella Borsa di Fabrizio, di "una "particolare formazione", costituita da una serie di lamine disposte a cipolla, con predominante struttura linforeticolare (vedi nota 8)". Questa "formazione" da una parte 'anticipa' la comparsa dei corpuscoli lamellari della Ghiandola Coccigea umana (Timo cloacale); dall'altra ne indica la derivazione e le trasformazioni (suggerendo anche 'l'origine' dei corpuscoli di Hassal del Timo). Tutti questi parallelismi tra Borsa di Fabrizio, Ghiandola Coccigea e principali ghiandole endocrine umane, non possono essere casuali: propongono per la Ghiandola Coccigea il ruolo di vera e propria ghiandola endocrina, deputata a regolare la caratterizzazione e la maturazione dei linfociti B (azione svolta dal Timo per i linfociti T).
Abbiamo visto che dalla Borsa di Fabrizio č stato isolato un fattore di maturazione dei linfociti B, la bursopoietina, ma che le ricerche sono state abbandonate perché questo peptide non si trovava nelle Borse di Fabrizio embrionarie (vedi nota 2). Sapendo che la Borsa di Fabrizio viene colonizzata dai linfociti B 6-8 giorni dopo che i linfociti T hanno colonizzato il Timo e che l'immunitŕ cellulo-mediata nel Pollo si matura circa una settimana dopo la nascita (vedi nota 6), č naturale che nelle Borse di Fabrizio embrionarie (non mature) la produzione di ormoni non sia dimostrabile.
Immunologia, Embriologia e Anatomia comparate suggeriscono che (vedi nota 26):
- nei Vertebrati inferiori, l'organo bursa-equivalente evolve dai ciechi rettali e dalla ghiandola digitiforme dei Pesci, ai ciechi dorsali degli Anfibi, alle ghiandole cloacali dei Rettili, alla Borsa di Fabrizio negli Uccelli, ai glomeruli caudali dei Mammiferi Inferiori;
- nel Pollo, la produzione dei fattori di maturazione e la maturazione stessa dei Linfociti B avvengono in un'unica sede anatomica, la Borsa di Fabrizio;
- nell'Uomo, con l'Evoluzione, si arriva alla separazione netta tra compiti e sedi: i fattori di maturazione dei Linfociti B sono secreti da una ghiandola endocrina specifica, la Ghiandola Coccigea, che nell'Evoluzione ha perso completamente le connessioni con l'Intestino Primitivo (ancora presenti in un Vertebrato inferiore, il Pollo); - la sede di maturazione č dislocata prima negli organi ematopoietici fetali, poi solo nel midollo osseo (in queste sedi, i Linfociti B sono anche protetti da interferenze di stimoli antigenici esterni).
Per conferma, credo sia doveroso il riesame anatomo-funzionale della Ghiandola Coccigea nell'Uomo; delle strutture anatomiche omologhe negli altri Vertebrati. In attesa delle necessarie verifiche: come organo deputato a regolare lo sviluppo e il perfezionamento dell'immunitŕ umorale, nell'Uomo, propongo la Ghiandola Coccigea.
Dr.
Domenico Fiore
medico libero professionista
Viale Madonna delle Grazie 17
35028 Piove di Sacco (PD)
Telefono: 049-9704306
www.domenicofiore.it
Note bibliografiche
1 - Dammacco F.:Immunologia in Medicina. Edi-Ermes. vol I, 2. 1989.
2 - Bach J.F.: Immunologie. Flammarion. Paris. 53-58, 87-88. 1986.
3 - Romer A.S.: Anatomia comparata dei Vertebrati. Piccin. Padova 1978.
4 - Playfair J.H.L.: L'Immunologia a colpo d'occhio. Medico e Paziente, VII, n° 2, 32. 1981.
5 - Pera L: Morfologia e sviluppo della Borsa di Fabrizio in Gallus domesticus. Arch. Ital. Anatom. ed Embriol. Vol LXIII, 407 e seguenti. 1958.
6 - Park B.H.- Good R.A.: Principi di Immunologia Moderna. Piccin Ed. Padova. 26-28. 1977.
7 - Vannini E.: Zoologia dei Vertebrati. UTET, Torino. 507-509 + figura 7.45. 1982
8 - Dulzetto F.: Anatomia Comparata dei Vertebrati. Edizioni Calderini. Bologna. 1600. 1968.
9 - Tuchmann-Duplessis H.: Embriologia Umana. UTET. Torino. 154-157. 1981.
10 - Romer A.S.: Anatomia comparata dei Vertebrati. Piccin. Padova. 486-489. 1978.
11 - Testut L.: Anatomia Umana. UTET, Torino. 538-539. 1943.
12 - Williams R.H.: Trattato di Endocrinologia.Piccin Ed. Padova. Vol I. 36-38. 1979.
13 - Patrono V.: Endocrinologia per la clinica. Il Pensiero Scientifico Ed.. Roma. 627-628. 1981.
14 - Beaumont A.- Cassier P.: Anatomia comparata dei Vertebrati. Ed. Ambrosiana. Milano1974.
15 - Hamilton-Boyd-Mossman: Embriologia Umana. Piccin Ed Padova 1977.
16 - Testut L.: Anatomia Umana. UTET, Torino. Voll IV-IX-X. 1943.
17 - Chiarugi G.: Istituzioni di Anatomia dell'Uomo. Soc Ed Libraria. Milano. Voll II e III. 1946.
18 - Anatomia del Gray. Zanichelli Ed. Bologna - Vol. II, 1450. 1994.
19 - Vannini E.: Zoologia dei Vertebrati. UTET, Torino. 438 + fig. 6.69 , 443 + fig 6.80. 1982.
20 - Dulzetto F.: Anatomia Comparata dei Vertebrati. Calderini. BO. 1549-1550, 1946. 1968.
21 - Dulzetto F.: Anatomia Comparata dei Vertebrati. Calderini. Bologna. 1946. 1968.
22 - Dulzetto F.: Anatomia Comparata dei Vertebrati. Calderini. Bologna. 1489-1492. 1968.
23 - Dulzetto F.: Anatomia Comparata dei Vertebrati. Calderini. Bologna. 246. 1968.
24 - Vannini E.: Zoologia dei Vertebrati. UTET, Torino. 170-171 + fig 3.91-. 1982
25 - Dulzetto F.: Anatomia Comparata dei Vertebrati. Calderini. Bologna 1454. 1968.
26 - Fiore D.: L'0rgano bursa-equivalente nell'Uomo e nei Vertebrati. Fiore Editore. Polverara (PD). 1981.
L'organo bursa-equivalente nell'Uomo
Immunitŕ umorale e ghiandola coccigea
Dalle
ricerche del Dr. Fiore emerge che, nell'Uomo, l'organo che presiede allo
sviluppo e alla "maturazione" dell'immunitŕ umorale, equivalente
alla Borsa di Fabrizio degli Uccelli![]() , č la Ghiandola
Coccigea
, č la Ghiandola
Coccigea![]() .
.
Rinviando per le premesse storiche, anatomiche e fisiologiche alle mie precedenti pubblicazioni sull'argomento (L'Organo Bursa-Equivalente nell'Uomo e nei Vertebrati - Ed. Fiore - 1981; L'Organo Bursa-Equivalente nell'Uomo e nei Vertebrati. Riapertura del dibattito e proposta di verifica.. Articolo pubblicato in: EOS. Vol XIX, 1999. n° 1, pag. 8-11.), per una definitiva collocazione anatomo-fisiologica della Ghiandola Coccigea (č o no l'Organo Bursa-Equivalente nell'Uomo?), propongo ora una facile verifica con il testo che segue.
La Borsa di Fabrizio (B.F.) per il suo aspetto č nota anche come "Timo cloacale". In un Pollo di 6 settimane č stata descritta «una particolare formazione, costituita da una serie di lamine disposte a cipolla, con predominante struttura linforeticolare. Le singole lamine, separate da uno spazio ricoperto da epitelio cilindrico monostratificato contenente cellule caliciformi, derivano da pieghe epiteliali della parete embrionale della B.F. e si formano prima della schiusa » [8]. La formazione di linfociti nella B.F. inizia 5 o 6 giorni dopo che nel Timo [6]. Da B.F. prelevate dopo la nascita, č stata isolata la bursopoietina [2], un peptide capace d'indurre l'espressione di alcuni antigeni di differenziazione dei linfociti B. La produzione di questo fattore suggerirebbe per la B.F. una funzione ormonale; ma le ricerche in questa direzione sono state abbandonate perché, utilizzando B.F. embrionarie, non si sono ottenuti risultati positivi. Dopo la nascita la B.F. subisce una involuzione direttamente collegata con la maturazione sessuale: comincia ad atrofizzarsi verso il 4° mese di vita (pubertŕ, nei Polli) ed č praticamente scomparsa attorno all'anno [2]. La B.F. č irrorata dall'Arteria Pudenda Communis [5], ramo dell'Arteria Sacrale Media.
Abbiamo visto che dalla B.F. č stato isolato un fattore di maturazione dei linfociti B, la bursopoietina, ma che le ricerche sono state abbandonate perché questo peptide non si trovava nelle B.F embrionarie [2]. Sapendo [6] che la B.F. viene colonizzata dai linfociti-B 6-8 giorni dopo che i linfociti T hanno colonizzato il Timo e che l'immunitŕ cellulo-mediata nel Pollo si matura circa una settimana dopo la nascita, č naturale che nelle B.F. embrionarie (non mature) la produzione di ormoni non sia dimostrabile.
La Bursopoietina si puň trovare (e si deve ricercare) solo nei Polli con almeno 15-20 giorni di vita: non si deve ricercare negli embrioni (nei quali non si puň trovare perché la Borsa di Fabrizio non č ancora "matura"); non si deve ricercare in Polli con piů di 4 (quattro) mesi di vita (la B.F. si atrofizza alla pubertŕ ed č praticamente scomparsa attorno all'anno di etŕ, nel Pollo).
Dopo la scoperta della funzione della Borsa di Fabrizio negli Uccelli (B. Glick 1956), anche nell'Uomo e negli altri Vertebrati si č ricercato affannosamente un organo con tale funzione:
Organo Bursa-Equivalente nell'Uomo
Sono stati studiati: tonsille, appendice ciecale (specie nel coniglio), placche di Peyer, G.A.L.T. (tessuto linfoide associato al tubo digerente), fegato, midollo osseo. Nessuna di queste formazioni ha caratteri di organo linfoide primario (centrale).
Si č arrivati alla conclusione che «nei Mammiferi non esiste un organo bursa-equivalente e la maturazione dei linfociti B avviene, negli stadi piů precoci dello sviluppo, a livello di organi emopoietici quali il fegato fetale, e, successivamente, nel midollo osseo [2, 3]».
Questa conclusione č in evidente contrasto con l'Anatomia e l'Immunologia Comparate. La Classificazione del Regno Animale [3], sovrapponibile alla Scala dell'Evoluzione, separa gli Invertebrati dai Vertebrati. Nei Vertebrati si passa: dai Pesci agli Anfibi; dagli Anfibi ai Rettili; da questi: da un lato agli Uccelli, dall'altro ai Mammiferi.
Negli Invertebrati non esistono organi linfoidi, ma solo cellule linfoidi libere. Nei Vertebrati, dalla Lampreda in su, č sempre presente il Timo: da masserelle irregolari poste dorsalmente alle fessure branchiali (Pesci) e da masse di tessuto lobulato diffuso (Anfibi) si passa a due cordoni ghiandolari ai lati del collo (Rettili), a due strisce di tessuto ghiandolare irregolarmente lobulate lungo tutto il collo (Uccelli) e infine all'organo pari (Mammiferi).
Seguendo lo sviluppo dell'immunitŕ umorale [4], vediamo che:
- nei Pesci: la Lampreda produce una immunoglobulina primitiva (formata da una molecola labile a 4 catene);
- nei Pesci cartilaginei (Squali) compaiono le plasmacellule e le catene Ig sono legate da ponti disolfuro, ma c'č ancora un'unica classe Ig;
- nei Teleostei si dimostra una primordiale cooperazione tra linfociti T e B, si differenziano gli Antigeni di I e II classe;
- negli Anfibi e nei Rettili compaiono i gangli linfatici e una nuova classe immunoglobulinica, le IgG;
- negli Uccelli compare improvvisamente (?!) la Borsa di Fabrizio e si ha dualitŕ netta delle linee linfocitarie T e B, ma nel Complemento mancano ancora C4 e C2;
- nei Mammiferi compaiono tutte le classi di immunoglobuline: «i Mammiferi sono caratterizzati piů dalla diversitŕ delle classi e sottoclassi di immunoglobuline che da un ulteriore sviluppo dei Linfociti T». Si avrebbe, nei Mammiferi, un perfezionamento della immunitŕ umorale senza il parallelo perfezionamento dell'Organo che negli Uccelli presiede alla "caratterizzazione" preliminare specifica dei Linfociti B.
Secondo Bach (Bach J-F: Ontogenesi delle Immunoglobuline nell'Uomo. Immunologie. Flammarion. Paris, 1986. 294-295) «La produzione delle immunoglobuline non ha raggiunto la sua maturitŕ alla nascita. Certamente, dalla fine della vita fetale esistono dei linfociti portatori di immunoglobuline di membrana, in particolare IgM e IgD, ma non č dimostrabile ancora alcuna produzione di IgG e il tasso delle IgM e delle IgA č ancora molto flebile. I tassi serici elevati di IgG notati nelle prime settimane di vita sono di origine materna.Le IgG non sono prodotte prima dell'etŕ da due a quattro mesi, cioč dopo la sparizione delle IgG materne. C'č dunque un periodo di qualche settimana verso l'etŕ di tre mesi in cui il tasso delle IgG seriche č molto basso. La produzione di IgM č la prima a raggiungere il livello osservato nell'adulto, seguita da quella delle IgG e con molto lungo intervallo da quella delle IgG e delle IgE. Č interessante notare che l'aumento della produzione di queste differenti immunoglobuline č influenzata dalle infezioni come lo dimostrano lo sviluppo ritardato e il livello ridotto della sintesi delle immunoglobuline negli animali axenici (germe-free) e al contrario l'apparizione anormalmente rapida della produzione dell'immunoglobuline nei bambini infettati in utero».
Secondo Male (Male D. et Altri: Immunoglobuline nel siero del fato e del neonato. Immunologia. Elsevier Masson. 2006. Pag. 76, figura 3.19) «Le IgG nel feto e nel neonato derivano unicamente dalla madre. Queste IgG materne scompaiono dall'etŕ di nove mesi, momento dal quale il neonato inizia a sintetizzare le proprie IgG. Il neonato produce le proprie IgM e IgA; queste classi non possono attraversare la placenta. dall'etŕ di 12 (dodici) mesi, il lattante produce l'80% del suo livello adulto di IgG, il 75% del suo livello adulto di IgM e il 20% del suo livello adulto di IgA».
In pratica, tenendo conto della produzione di immunoglobuline nel Feto e nel Neonato (Bach J-F; Male et Altri) la bursopoietina si puň trovare nella Ghiandola Coccigea (G.C.) in bambini deceduti all'etŕ di 8-12 mesi: non molto prima, perché la G.C. non č ancora "matura"; non molto dopo, per l'involuzione fisiologica della ghiandola (vedere l'involuzione della B.F. nel Pollo e del Timo nell'Uomo).
Nell'Uomo: tenendo presente l'ontogenesi delle immunoglobuline, la bursopoietina si deve ricercare nella Ghiandola Coccigea. Nell'Uomo [9] il Timo produce diversi ormoni dotati di attivitŕ biologica certa:
- la Timosina (induce la differenziazione dei linfociti-T; aumento della responsivitŕ dei linfociti ai mitogeni policlonali; aumento delle cellule T-helper);
- la Timopoietina (interferisce con la trasmissione neuromuscolare; induce la maturazione dei linfociti da midollari a linfociti intratimici);
- la Timostimolina (promuove la maturazione dei linfociti);
- la Timomodulina (attiva le cellule immunocompetenti):
- il Fattore timico X ( attiva le cellule immunocompetenti).
Per analogia con quanto avviene nel Timo, nella Ghiandola Coccigea e nella Borsa di Fabrizio, oltre alla Bursopoietinasi potranno trovare (e si dovranno ricercare) altri fattori ormonali coinvolti nella differenziazione dei Linfociti.
Dr.
Domenico Fiore
medico libero professionista
Viale Madonna delle Grazie 17
35028 Piove di Sacco (PD)
Telefono: 049-9704306
www.domenicofiore.it
Bibliografia
2) Bach J.F.: Immunologie. Flammarion. Paris. 53-58, 87-88. 1986.
3) Romer A.S.: Anatomia comparata dei Vertebrati. Piccin. Padova 1978.
4) Playfair J.H.L.: L'Immunologia a colpo d'occhio. Medico e Paziente, VII, n° 2, 32. 1981.
5) Pera L: Morfologia e sviluppo della Borsa di Fabrizio in Gallus domesticus. Arch. Ital. Anatom. ed Embriol. Vol LXIII, 407 e seguenti. 1958.
6) Park B.H.- Good R.A.: Principi di Immunologia Moderna. Piccin Ed. Padova. 26-28. 1977.
7) Vannini E.: Zoologia dei Vertebrati. UTET, Torino. 507-509 + figura 7.45. 1982
8) Dulzetto F.: Anatomia Comparata dei Vertebrati. Edizioni Calderini. Bologna. 1600. 1968.
9) Indiveri F.: Ormoni Timici. In Dammacco: Immunologia in Medicina. edi-ermes.Milano 1989. 1377-1389.
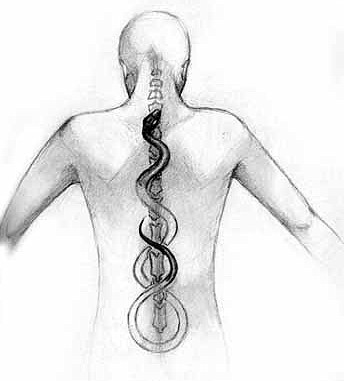
Una ghiandola per la Kundalini
Quante altre
ghiandole minuscole attendono di essere conosciute? E quante ancora, quali
ricettori non usati, sono presenti nel nostro corpo come potenziale nascosto e
inutilizzato, e quindi pronte a prender forma? La Kundalini![]() allora non
appare piů alla scienza come un vocabolo newage per energetici incalliti e
beoti dello spirito, ma ha una sua specificitŕ fisio-anatomica che
corrisponde a una ghiandola particolare: il glomo coccigeo del Luschka
allora non
appare piů alla scienza come un vocabolo newage per energetici incalliti e
beoti dello spirito, ma ha una sua specificitŕ fisio-anatomica che
corrisponde a una ghiandola particolare: il glomo coccigeo del Luschka![]() .
.
Il seguente intervento, parte di un convegno della serie: la scienza incontra lo spirito - Sessualitŕ e spiritualitŕ č ascoltabile integralmente su dvd dal cofanetto che prende lo stesso nome. Alla fine dell'articolo avete l'occasione di visionarne il trailer.
La Ghiandola
Coccigea del Luschka, identificata per la prima volta a metŕ dell'ottocento
dal famoso anatomo-patologo dell'epoca Hubert Luschka![]() di Tübingen
(1820–1875), č un piccolo granellino di 3-4 millimetri che si trova davanti
al coccige. Ha dei canali vascolari permeati di fibre nervose e sinusoidi
vascolari, con ampia innervazione. Dalla struttura anatomica e istologica
sembrerebbe un chemocettore.
di Tübingen
(1820–1875), č un piccolo granellino di 3-4 millimetri che si trova davanti
al coccige. Ha dei canali vascolari permeati di fibre nervose e sinusoidi
vascolari, con ampia innervazione. Dalla struttura anatomica e istologica
sembrerebbe un chemocettore.
Dei
chemocettori conosciuti, quali il glomo carotideo e il glomo aortico, si
apprendono le importantissime funzioni come quella di rilevare la pressione
parziale di ossigeno e di anidride carbonica nel sangue, informando
direttamente il cervello e il midollo allungato, al fine di dare la risposta
respiratoria adeguata. Anche se, purtroppo, non ci sono grandi studi su questa
parte anatomica, gli esperimenti da noi condotti hanno avuto riscontri
importantissimi. Considerando che, l'inizio della catena del “simpatico”
č la Ghiandola Pineale o epifisi![]() (importantissima sul controllo dell'immunitŕ) e questo glomo coccigeo č la
parte finale, si arriva per deduzione a supporre il ruolo di questo organo sul
controllo dell'identitŕ biologica, la quale deve corrispondere a una identitŕ
psicobiofisica (o sessuale) e a una spirituale, anche se non vi č ancora una
coscienza di ciň. Asportando il glomo del Luschka diminuisce la capacitŕ
biologica di riconoscere la propria identitŕ biologica.
(importantissima sul controllo dell'immunitŕ) e questo glomo coccigeo č la
parte finale, si arriva per deduzione a supporre il ruolo di questo organo sul
controllo dell'identitŕ biologica, la quale deve corrispondere a una identitŕ
psicobiofisica (o sessuale) e a una spirituale, anche se non vi č ancora una
coscienza di ciň. Asportando il glomo del Luschka diminuisce la capacitŕ
biologica di riconoscere la propria identitŕ biologica.
Apro inoltre una parentesi per dire che la stimolazione della Ghiandola Coccigea (o glomo del Luschka) puň avvenire anche durante il rapporto anale, il cui piacere č pertanto semplicemente dovuto alla stimolazione della stessa e non alla penetrazione anale in sé.
Nella tradizione dei chakra, la Ghiandola Pineale č connessa all'espansione della coscienza e allo Spirito, e corrisponde al settimo chakra, Brahmarandhra o Loto dai Mille Petali; la Ghiandola Coccigea, che č legata alla sessualitŕ e al piacere, corrisponde quindi al primo chakra Mulhadara o Kundalini, e viene messa in comunicazione con la Pineale attraverso il “simpatico”. Se tale relazione fra queste due strutture divenisse cosciente, vi sarebbe davvero la possibilitŕ di riportare la sessualitŕ da fatto puramente materiale a fatto celeste.
Fernando
Brivio
chirurgo, ricercatore, teologo e docente di PNEI
www.scienzaeconoscenza.it
![]()
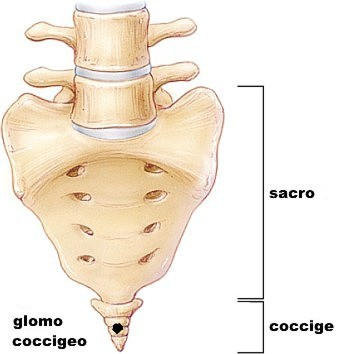
Si trova, nell’uomo, direttamente applicata sulla
faccia anteriore dell’ultimo pezzo del coccige, un piccolo corpo
tondeggiante o lobulato, avente appena qualche millimetro di diametro: č la ghiandola
coccigea del Luschka![]() (glomus
coccygeus).
Nonostante il suo nome, questa formazione non presenta una struttura
ghiandolare: secondo alcuni (Gegenbaur, Cunéo e Veau), essa rappresenterebbe
la porzione terminale dell’arteria sacrale media, la quale, primitivamente,
avrebbe irrigato le vertebre caudali scomparse attualmente nell’uomo;
secondo altri (Jakobson, 1899), sarebbe il vestigio della porzione caudale del
simpatico.
(glomus
coccygeus).
Nonostante il suo nome, questa formazione non presenta una struttura
ghiandolare: secondo alcuni (Gegenbaur, Cunéo e Veau), essa rappresenterebbe
la porzione terminale dell’arteria sacrale media, la quale, primitivamente,
avrebbe irrigato le vertebre caudali scomparse attualmente nell’uomo;
secondo altri (Jakobson, 1899), sarebbe il vestigio della porzione caudale del
simpatico.
Checché sia del suo vero significato, non sembra che il glomo coccigeo abbia nella patogenesi dei tumori congeniti sacro-coccigei l’importanza a esso attribuita da Braune, Virchow, Perrin, Sertoli, Lachaud (citati dal Rieffel). La maggior parte di questi tumori, giŕ lo dicemmo, sono consecutivi, sia a una malformazione dell’estremitŕ inferiore del canale sacrale, sia a una proliferazione di elementi cellulari che rappresentano il vestigio della midolla coccigea (Tourneux ed Hermann) e, forse, ai resti dell’intestino caudale dell’embrione.
Il coccige č un osso impari e
simmetrico, ultimo verso il basso della colonna vertebrale. Deriva il suo nome
dal greco kókkyx, propriamente "cuculo", per via della
somiglianza con il becco di tale uccello. Il primo a utilizzare il termine con
tale significato fu il medico Galeno![]() nel II secolo dC. Il coccige č l’ultimo segmento rudimentale della colonna
vertebrale ed č formato da 3 a 5 segmenti fusi tra loro che perň non
presentano le normali caratteristiche delle vertebre ma sono, al contrario,
soltanto abbozzi di queste ultime. Solo la prima vertebra coccigea presenta
ancora qualche caratteristica comune alle vertebre precedenti: si notano
infatti gli abbozzi dei processi trasversi e dei processi articolari, qui
definiti corni del coccige, situati posteriormente alla base e che si
articolano con i corni del sacro.
nel II secolo dC. Il coccige č l’ultimo segmento rudimentale della colonna
vertebrale ed č formato da 3 a 5 segmenti fusi tra loro che perň non
presentano le normali caratteristiche delle vertebre ma sono, al contrario,
soltanto abbozzi di queste ultime. Solo la prima vertebra coccigea presenta
ancora qualche caratteristica comune alle vertebre precedenti: si notano
infatti gli abbozzi dei processi trasversi e dei processi articolari, qui
definiti corni del coccige, situati posteriormente alla base e che si
articolano con i corni del sacro.
Nell’insieme l’osso ha forma triangolare con una base superiore, un vertice inferiore, una faccia anteriore, una posteriore e due margini laterali. La base si articola con l’apice del sacro. Le facce anteriore e posteriore presentano dei solchi orizzontali che segnano i punti di fusione dei segmenti primitivi. Č l'ultima testimonianza della coda che caratterizzava la nostra specie in epoca remota. A prova di ciň si riscontra, anche se molto raramente, in alcuni nascituri la presenza di ulteriori vertebre (generalmente 2-3 ma anche 5) al di sotto di esso che formano una "piccola coda". Sulla superficie dorsale del coccige si inserisce il grande gluteo, verso l'apice del coccige si inserisce invece lo sfintere esterno dell'ano.
Testut
& Jacob
Anatomia topografica
UTET - 1960
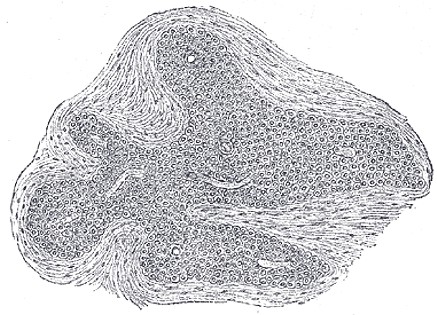
Section of an irregular nodule of the glomus coccygeus x 85
The section shows the fibrous covering of the nodule, the bloodvessels within it, and the epithelial cells of which it is constituted. (Henry Gray, 1918)
The coccygeal glomus (coccygeal gland or body; Luschka’s gland, Latin glomus coccygeus) is placed in front of, or immediately below, the tip of the coccyx. It is about 2.5 mm. in diameter and is irregularly oval in shape; several smaller nodules are found around or near the main mass.
It consists of irregular masses of round or polyhedral cells, the cells of each mass being grouped around a dilated sinusoidal capillary vessel. Each cell contains a large round or oval nucleus, the protoplasm surrounding which is clear, and is not stained by chromic salts.

![]()
Centro in provincia di Viterbo, 50 km a NW del capoluogo, a 420 m su uno sprone dominante la valle del fiume Paglia. Comune di 130,28 kmq con 5868 abitanti. Agricoltura (cereali, uva); allevamento bovino e avicolo; industrie del mobile e dei laterizi. Di probabile origine etrusca (forse l'antica Acula), fu fortificata nel Medioevo pervenendo nel 1115 alla Santa Sede e passando successivamente al Comune di Siena e agli Sforza. Ebbe statuti propri dal 1443; dopo la distruzione di Castro (1649) divenne sede vescovile e rimase definitivamente soggetta alla Chiesa. La cattedrale, basilica benedettina romanica piů volte restaurata, č caratterizzata da un impianto a croce latina e tre absidi; nell'interno, conserva bassorilievi di Agostino di Duccio. La cripta (sec. XI), a nove navate, ha le pareti decorate con resti di affreschi del sec. XIII; nel Medioevo era meta di pellegrinaggio per i Crociati e le genti dirette in Terra Santa.
Acquapendente
Stato:Italia
Regione:Lazio
Provincia:Viterbo
Altitudine: 420 m s.l.m.
Superficie: 120,28 km˛
Abitanti: 5.776 (2007)
Densitŕ: 48,02 ab./km˛
Frazioni: Torre Alfina, Trevinano
Nome abitanti: aquesiani
Santo patrono: Sant'Ermete
Giorno festivo: 28 agosto
Acquapendente č un comune di 5.776 abitanti in provincia di Viterbo posto dieci chilometri a nord del Lago di Bolsena, presso la Riserva naturale Monte Rufeno che si trova sulla riva opposta del fiume Paglia; la cittadina č raggiunta dalla Via Cassia. Acquapendente deve il suo nome al fatto di essere situata nei pressi di numerose piccole cascatelle che confluiscono nel piccolo fiume Paglia, che segna il confine naturale tra Lazio e Toscana, sebbene il confine sia posto a qualche chilometro a nord del fiume.

Il paese č stato storicamente per lungo tempo sotto il dominio di Siena, sebbene formalmente fosse stato lasciato in ereditŕ alla Chiesa dalla contessa Matilde di Canossa nel XII secolo. Nel XVI secolo il paese tornň sotto il dominio della Chiesa e papa Gregorio XIII fece costruire un ponte sul fiume, i cui piloni costruiti dall'architetto Domenico Fontana sono ancora quelli che sostengono il ponte attuale. Dalla valle formata dal fiume Paglia č possibile vedere in lontananza il Monte Amiata, che con i suoi 1738 m č la vetta piů alta della Toscana meridionale.
Nella seconda metŕ del 1600 Acquapendente visse un periodo d'intenso sviluppo, dopo la distruzione del ducato di Castro avvenuta nel 1649. Papa Innocenzo X fece distruggere la cittŕ di Castro per ridurre l'importanza della famiglia Farnese nella parte settentrionale del Lazio e fece poi spostare ad Acquapendente la sede del vescovo di Castro. La nuova situazione fece aumentare l'importanza della cittadina all'interno dello Stato Pontificio. Questo spiega anche il motivo per cui il papa commissionň ad Alessandro Algardi la realizzazione di una statua rappresentante il busto del papa e la fece mandare ad Acquapendente dove č ora conservata all'interno della cattedrale. Acquapendente fu testimone nel 1867 della Campagna risorgimentale dell'Agro Romano per la liberazione di Roma.
Nella cattedrale č conservata una pietra segnata con del sangue che si dice provenga dal Santo Sepolcro di Gerusalemme. La reliquia č conservata all'interno di una cripta di stile gotico molto interessante. La cattedrale č stata oggetto nel corso dei secoli di numerosi interventi di abbellimento; in particolare ci sono molte opere in terracotta dipinta.
Acquapendente nel medioevo era circondata da una cinta muraria della quale sfortunatamente non resta molto perché fu distrutta nei momenti di espansione della cittadina; resta comunque una delle torri principali, recentemente ristrutturata. La chiesa di San Francesco č una chiesa precedente alla nascita del santo e a lui dedicata successivamente. Il campanile č di epoca rinascimentale, ma il portale risale all'XI secolo. L'influenza degli artisti toscani č evidente nella decorazione a bugnato di Palazzo Viscontini, un bel palazzo rinascimentale con inusuali grandi finestre.
La terza domenica di maggio si svolge la Festa dei Pugnaloni per celebrare la Festa della Madonna del Fiore. Comprende eventi folkloristici e sfilate di mosaici realizzati con fiori e foglie.
Acquapendente

Fa parte dell' Alta Tuscia (o Etruria, anticamente abitata dagli Etruschi), ai confini con Toscana e Umbria. Prima del 1964, quando la via consolare Cassia rappresentava il collegamento preferenziale tra Firenze e Roma, Acquapendente era al centro di un consistente movimento che sosteneva l'economia di ristoranti (alcuni dei quali diventati famosi), alberghi e locande. Nei secoli passati la cittadina costituiva anche una statio privilegiata nel cammino dei pellegrini che, attraverso la Via Francigena, si dirigevano a Roma e alle coste della Puglia verso la Terra Santa.
Il primo documento storico ci porta al 964, quando Oddone I vi costruě un castello che poi passň agli Svevi. Il monumento piů rappresentativo č la cattedrale del Santo Sepolcro, sorta intorno all'anno Mille sul sacello che Matilde di Westfalia fece erigere con le stesse dimensioni del Sepolcro di Cristo. Consacrata nel 1149 e piů volte ristrutturata, acquistň nel 1649 il titolo di cattedrale. Dopo i danni dell'ultima guerra č stata in gran parte riedificata su progetto di Vincenzo Fasolo. Nell'interno, pregevole l'ancona (trasformazione di icona, immagine, pannello dipinto o scolpito da porsi sull'altare) nel transetto destro, opera in terracotta smaltata di Jacopo Beneventano (1522) raffigurante il Padreterno adorato dagli angeli; il coro ligneo (1685-1688) nell'abside č riferito all'artista Matteo Tedesco. Nella cappella di Sant' Ermete si venera il santo patrono, raffigurato in una statua di cera. Sotto il presbiterio, tra le due scale di accesso, č collocato un pregevole fonte battesimale del Trecento. La cripta risale alla seconda metŕ del X secolo: 9 navate con 22 colonne sormontate da ricchi capitelli. Venne eretta sulla primitiva edicola del Santo Sepolcro che si trova al centro, coperta da una piramide a base rettangolare. Secondo la tradizione custodisce una reliquia litica bagnata del sangue di Cristo.
La torre Julia di Jacopo, nelle adiacenze della chiesa, č quanto resta della rocca di Arrigo IV (oggi destinata a centro culturale e di accoglienza). La torre dell'Orologio (detta anche di Barbarossa), nella parte piů alta del paese, faceva parte del castello imperiale, a lungo tenuto dagli Svevi. Notevole il palazzo Comunale (grande portico neoclassico ); il monumento al centro della piazza ricorda Fabrizio d'Acquapendente (1533?-1619), nativo del posto e famoso per gli studi di anatomia.
Acquapendente č punto di riferimento per escursioni nella riserva naturale di monte Rufeno, che si estende nei dintorni: un'oasi di rilevante interesse ecologico attrezzata per escursioni e trekking. La riserva occupa circa 3000 ettari di terreni prevalentemente boschivi e in posizione collinare. Da quota 200 metri (fiume Paglia) si sale ai 700 metri del monte Rufeno.
Alcuni casolari offrono posti letto e ristoro con prodotti tipici: uno in particolare (il casale Giardino) ospita il museo del Fiore, con sezioni tematiche (introduzione al mondo del fiore, i fiori nella riserva di monte Rufeno, la morfologia del fiore, insetti e fiori, le curiositŕ nei fiori, i fiori e l'uomo) che utilizzano illustrazioni, fotografie e pannelli, plastici e strumenti informatici. L’ampio giardino č disponibile anche per colazioni al sacco.
I Pugnaloni - Secondo la tradizione, l'origine della festa risale al 1166, quando Acquapendente era governata da un tiranno del Barbarossa .«Č piů facile che quel ciliegio ormai secco da anni fiorisca - avrebbero amaramente commentato due contadini - che il nostro paese possa liberarsi dall'invasore». L'improvvisa e miracolosa fioritura dell'albero venne accolta come un segno della Madonna che diede agli acquesiani la forza e l'entusiasmo per la rivolta. In ricordo di quell'evento, un tempo si infiocchettavano i pungoli (bastoni di legno usati per guidare i buoi e combattere contro il nemico), che oggi sono diventati “Pugnaloni”, enormi mosaici in gran parte composti di fiori con immagini sulla libertŕ. Nel tardo pomeriggio della domenica successiva al 15 maggio vengono portati in processione dalla Cattedrale del Santo Sepolcro fino al palazzo Comunale.
I Pugnaloni (una ventina) vengono realizzati la notte precedente da gruppi di giovani divisi in quartieri; dopo la processione si procede al non facile compito dell’assegnazione dei premi alle migliori composizioni. Da qualche anno si č affermata anche la tradizione dei “ mini-pugnaloni” eseguiti con altrettanta bravura dai ragazzi del posto.
www.latuscia.com
Acquapendente
Situata sul margine di un ripiano che scende a nordovest nella valle del Paglia, č una vivace cittadina che risente degli influssi della vicina Toscana e comune agricolo fra i piů importanti del Viterbese con notevole produzione di legumi e vino e distretto artigianale con le sue fabbriche di ceramica. Le sue antiche origini risalgono ai Fallisci, quando si chiamava Acula, Aquila o Aquesium mentre in epoca romana divenne una ‘mansio’. Fortezza degli imperatori tedeschi degli Ottoni, [č ricordata la prima volta nel 964 come castello imperiale di Ottone I], passň poi agli Svevi e quindi entrň a far parte del marchesato di Toscana; attraverso l’ereditŕ di Matilde di Canossa pervenne alla Santa Sede, a cui perň fu contesa dal Barbarossa, finché nel 1166 si liberň dalla soggezione imperiale. Sede dell’Abbazia del Santo Sepolcro, i cui frati ebbero giurisdizione nella zona, partecipň alle crociate. Tra la fine del XIV e l’inizio del XV appartenne per venticinque anni al Comune di Siena. Successivamente dal 1415 vi dominarono gli Sforza. Consolidata la libertŕ cittadina [1143] con il capitolato di Eugenio IV, e fissato il suo jus municipale in uno statuto del tempo di Nicolň V, la cittadina, divenuta sede di diocesi sotto il papato di Innocenzo X dopo la distruzione di Castro [1649] restň in diretto dominio della Chiesa.
La prima domenica dopo il 15 maggio vi si tiene, a ricordo della cacciata del Barbarossa, la festa della Madonna del Fiore o di ‘Mezzo Maggio’ in cui vengono fatti sfilare i cosiddetti pugnaloni, grandi quadri a soggetti religiosi o allegorici, sempre variati, composti con foglie e petali di fiori. All’ingresso dell’abitato s’incontrano subito a sinistra la torre medievale detta ‘Julia de Jacopo’, porta delle antiche mura, che ospita al suo interno un Museo della Ceramica e a destra la Cattedrale di San Sepolcro, basilica consacrata nel 1149 dal vescovo Aldobrandino da Orvieto; costruita accanto all’Abbazia benedettina del Santo Sepolcro, documentata dal 1025, č incerto se vi fu annessa una casa di Cavalieri Templari. Rimaneggiata nella parte superiore verso la metŕ del 1700 nelle tipiche forme laziali dell’epoca, in seguito ai danneggiamenti subiti durante la seconda guerra mondiale č stata in parte ripristinata nelle antiche forme romaniche. L’imponente facciata č fiancheggiata da due campanili, e lungo il fianco destro corre un portico nel quale sono collocati resti dell’edificio romanico; delle tre absidi solo quella centrale č originaria. Il grandioso interno č a croce latina con tre navate divise da pilastri, col transetto e l’abside sopraelevati e copertura a capriate. Sotto il presbiterio di estende la grandiosa cripta del secolo IX adorna di affreschi del secolo XIII; č a nove navate divise da basse colonne con ricchi capitelli romanici reggenti volte a crociera gotiche; in una cappella a sinistra, entro un altare retto da un cippo pagano, sono le reliquie ritenute della colonna della Flagellazione. A un piano inferiore č un sacello che custodisce le reliquie ritenute dell’Aula del Pretorio di Gerusalemme: le dimensioni e l’orientamento del sacello sono quelle del Santo Sepolcro di Gerusalemme, dal quale il titolo della chiesa frequentata dagli antichi pellegrini di Terrasanta e dei Crociati.
Uscendo dalla chiesa, di fronte, sopra un colle, seminascosto da una piccola pineta, č visibile la Torre dell’Orologio, detta anche del Barbarossa, resto del castello imperiale probabilmente del XII secolo. Da qui si segue la Via Roma rasentando il fianco della chiesa di Sant'Agostino e si continua nella Via Battisti, dove s’incontra al civico 35 il Palazzo Viscontini di Ippolito Scalza, con ricco portale a bugne e un piano di finestre architravate. Poco dopo č la piazzetta con la chiesa di San Francesco precedentemente intitolata a Santa Maria, originariamente gotica, dal bel portale trilobato con tracce di protiro e campanile cinquecentesco isolato a due piani di monofore e uno di bifore. Usciti dalla chiesa si retrocede e si volge a destra nel corso Taurelli Salimbeni che porta alla vasta Piazza Comunale dove sono situati il monumento di Fabrizio d’Acquapendente di Tito Sarrocchi [1888] e il neoclassico Palazzo Comunale, a portico terreno. Di fronte sorge la chiesa di Santo Stefano, rifatta dopo la seconda guerra mondiale sulle rovine della precedente chiesa seicentesca.
www.tusciaromana.it
Acquapendente is a city in the province of Viterbo, in Lazio (Italy). Acquapendente is an important centre for agricultural production of legumes and wine, and has a tradition in pottery craftmanship. The area of modern Acquapendente was settled by Etruscans and Roman times, as findings have showed. However, the first historical document of the modern city date from the 9th century AD, with a burg named Farisa or Arisa along the Via Francigena. In 964 a document of Emperor Otto I contains for the first time the name Aquapendentem. The name of the city, meaning "Pending Water", stems from the presence of several small waterfalls forming the Paglia, a stream setting the boundary between Lazio and Tuscany.
The city was later part of the March of Tuscany and, from the end of the 14th century and beginning of the 15th, it was part of the commune of Siena. In 1449 it become an independent centre within the Papal States- The Diocese of Acquapendente was under the ecclesiastical jurisdiction of Orvieto until 1649, after the War of Castro. Acquapendente was also the birthplace of the anatomist Hieronymus Fabricius.
![]()
Hubert von Luschka, born Hubert Luschka (Konstanz July 27, 1820 – Tübingen March 1, 1875), was a German anatomist. He lent his name to several structures, including the foramina of Luschka, Luschka's crypts, Luschka's joints, coccygeal Luschka’s gland and ducts of Luschka. He began studying medicine, initially Pharmacology, in 1841 at the University of Freiburg and the University of Heidelberg. In 1845 he became an assistant to Louis Stromeyer in Freiburg, then moved in 1849 to the University of Tübingen, where he was a lecturer and extraordinary professor until being appointed regular Professor of Anatomy in 1855. He gained a noble title and began using von in his name in 1865.
His work particularly concerned the need for anatomy to be connected in a practical manner to medicine and surgery. His Anatomie des Menschen in Rücksicht auf das Bedürfnis der praktischen Heilkunde (1862–69; Human Anatomy in Consideration of the Needs of Practical Medicine) aimed to provide such a link. He promoted the use of anatomical information in surgery, for example to manipulate internal organs using long needles before cutting the body open, and was one of the first to conduct detailed research on normal corpses (rather than only diseased or anomalous ones), publishing a series of detailed books covering specific aspects of anatomy, such as the nerves of the hands and the blood vessels of the brain.
Works
Die Nerven in der harten Stirnhaut
(1850)
Die Struktur der serösen Häute des Menschen (1851)
Der nervus phrenicus des Menschen (1853)
Die Adergeflechte des menschlichen Gehirns (1855)
Die Brustorgane des Menschen in ihrer Lage (1857)
Die Halbgelenke des menschlichen Körpers (1858)
Die Halsrippen und die ossa suprasternalia (1859)
Der Herzbeutel und die Fascia endothoracica (1859)
Der Hirnanhang und die Steißdrüse des Menschen (1860)
Anatomie des Menschen in Rücksicht auf das Bedürfnis der praktischen
Heilkunde (1862–69)
Der Schlundkopf des Menschen (1868)
Über Maß- und Zahlenverhältnisse des menschlichen Körpers (1871)
Der Kehlkopf des Menschen (1871)
Die Lage der Bauchorgane (1873)
Hubert von Luschka (* 27. Juli 1820 in Konstanz; † 1. März 1875 in Tübingen) war ein deutscher Anatom. Hubert von Luschka beschäftigte sich zunächst mit Pharmazie, studierte Medizin ab 1841 an der Universität Freiburg, 1843/1844 an die Universität Heidelberg. Sein Staatsexamen machte er 1844 in Karlsruhe, seine Praxiszeit absolvierte er in Meersburg. 1845 wurde er in Freiburg promoviert.
Er absolvierte eine Assistenz bei Louis Georg Friedrich Stromeyer (1804–1876), unternahm Forschungsreisen nach Paris, Venedig und Norditalien und war als Arzt in seiner Heimatstadt Konstanz tätig.1849 erhielt er einen Ruf als außerordentlicher Professor für Anatomie an die Universität Tübingen. Als Nachfolger von Friedrich Arnold (1803–1890), der nach Heidelberg wechselte, wurde er 1852 Professor für Anatomie und Pathologie, ab 1855 Ordinarius für Pathologie und Direktor des Instituts für Anatomie. Er blieb in dieser Stellung bis zu seinem Tode 1875. Sein Nachfolger wurde Wilhelm Henke (1834–1896).
Luschka war einer der renommiertesten Anatomen des 19. Jahrhunderts. Er forschte über fast alle Bereiche des menschlichen Körpers und veröffentlichte zahlreiche wissenschaftliche Arbeiten. Hubert von Luschka veröffentlicht 1862 bis 1869 bei H. Laupp’schen Buchhandlung in Heidelberg eine mit zahlreichen Holzschnitten sehr aufwendig gestaltete Anatomie des Menschen in Rücksicht auf die Bedürfnisse der praktischen Heilkunde. Seinen Namen tragen heute noch einige anatomische Strukturen, z. B. ein Abfluss einer Hirnkammer sowie der Luschka’sche Gang.
Schriften
Entwicklungsgeschichte der
Formbestandtheile des Eiters und der Granulationen. Dissertation Freiburg,
1845.
Die Nerven in der harten Hirnhaut. Tübingen, 1850.
Die Nerven des menschlichen Wirbelcanals. Tübingen, 1850.
Die Structur der serösen Häute des Menschen. Tübingen, 1851.
Der Nervus phrenicus des Menschen. Tübingen, 1853.
Die Adergeflechte des menschlichen Gehirns. Berlin, 1855.
Die Brustorgane des Menschen in ihrer Lage. Tübingen, 1857.
Die Halbgelenke des menschlichen Körpers. Berlin, 1858.
Der Herzbeutel und die Fascia endothoracica. Denkschriften der kaiserlichen
Akademie der Wissenschaften in Wien, 1859.
Die Blutgefässe der Klappen des menschlichen Herzens. Denkschriften der
kaiserlichen Akademie der Wissenschaften in Wien, 1859.
Der Hirnanhang und die Steissdrüse des menschlichen Herzens. Berlin, 1860.
Die Muskulatur am Boden des weiblichen Beckens. Denkschriften der kaiserlichen
Akademie der Wissenschaften in Wien, 1861.
Ueber polypöse Vegetationen der gesammten Dickdarmschleimhaut. Virchows
Archiv für pathologische Anatomie und Physiologie und für klinische Medizin,
Berlin, 1861, 20: 133-142.
Die Anatomie des Menschen in Rücksicht auf die Bedürfnisse der praktischen
Heilkunde bearbeitet. 3 Bde., Tübingen, 1862–1867.
Die Anatomie des menschlichen Bauches/Die Anatomie des menschlichen Beckens.
Laupp, Tübingen 1863–1864.
Die Anatomie des menschlichen Halses/Die Anatomie der Brust des Menschen.
Laupp, Tübingen 1862–1863.
Die Venen des menschlichen Halses. Denkschriften der kaiserlichen Akademie der
Wissenschaften in Wien, 1862.
Der Schlundkopf des Menschen. Denkschriften der kaiserlichen Akademie der
Wissenschaften in Wien, 1868.
Die Lage der Bauchorgane des Menschen. 1873.
Der Kehlkopf des Menschen. Laupp, Tübingen, 1871.

Kundalini
Rising
Oil and Plaka in the collection of Celeste McNeal
Tulsa – Oklahoma – USA
Kundalini č un termine della filosofia Advaita e di altre tradizioni yogiche tra le quali quella classica di Patanjali. Con Kundalini si intende un'energia che risiede nel corpo umano a livello sottile, manifestazione dell'energia universale meglio nota come shakti. In particolare Kundalini corrisponderebbe alla "forza generativa" in contrapposizione alle altre due forme di energia tradizionali cioč prana (o energia vitale) e fohat (o energia di movimento).
Kundalini, allo stato latente, risiede alla base della colonna vertebrale sopra un plesso che si fa corrispondere al piů basso dei chakra, detto in genere mulādhāra, e in particolare nell'osso sacro. Essa č tradizionalmente rappresentata da un serpente addormentato, avvolto intorno alla base della spina dorsale in tre giri e mezzo. Il suo nome deriva dalla parola kundala, che significa avvolto, arrotolato, spiraliforme.
Fin dall'antichitŕ, il serpente č stato
considerato simbolo di trasformazione, grazie alla sua capacitŕ di mutare la
pelle ed č stato associato al benessere fisico, spirituale e
all'illuminazione. Anche il Bastone di Asclepio![]() o Caduceo di Ermete, simbolo della moderna medicina, ci presenta un serpente
che si avvolge attorno a un bastone.
o Caduceo di Ermete, simbolo della moderna medicina, ci presenta un serpente
che si avvolge attorno a un bastone.
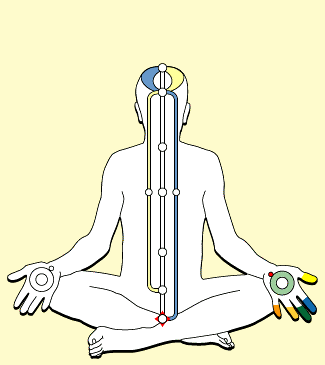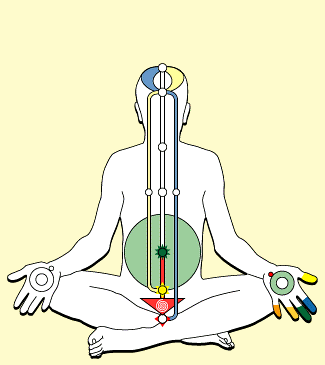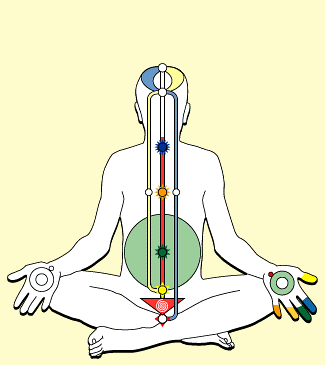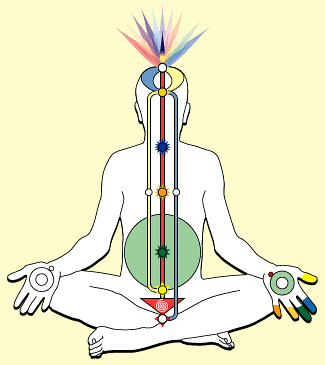
Kundalini č luce, č come l'intenso bagliore di una giovane e possente folgore, č suono, č Mantra, il suo dolce sussurro č simile al confuso ronzio di sciami di api in amore, essa produce melodiosa poesia. Secondo gli yogin, tale energia va risvegliata e dinamizzata attraverso la sadhana (pratica della disciplina) e spinta entro la direttrice centrale, detta Susumna o Shusumna, nella fisiologia sottile delle tre principali nadi del corpo umano, cosě da risalire i chakra superiori fino alla sommitŕ del cranio.Lungo il suo percorso, Kundalini attraversa i chakra risvegliandoli e purificandoli. Giunta al settimo chakra, detto Sahashrara o anche "loto dai mille (petali)", essa completa il suo risveglio, portando l'individuo nello stato che viene comunemente definito realizzazione del sé o risveglio secondo la tradizione indiana.
L'innalzarsi di questa energia permetterebbe di sperimentare, in piena consapevolezza, nuovi stati di coscienza e sarebbe solitamente accompagnato da una serie di fenomeni particolari che possono essere di tipo fisico, sensoriale, percettivo. L'effettiva manifestazione del risveglio della Kundalini č la percezione di aria, di un leggero soffio, di un leggero vento, di una brezza fresca sulla sommitŕ della testa e sui palmi delle mani.
![]()