Vol. 3° - X.
GENI attivi SU FUNZIONI E STRUTTURE
|
|
1. MUSCOLATURA
La muscolatura influenza per circa il 40% il peso corporeo.
Dobbiamo anche ricordare che il 40% delle proteine che compongono l’organismo
sono presenti nel tessuto muscolare. Sarebbe quasi superfluo far rilevare che
lo sviluppo delle masse muscolari dei broilers assume una notevole importanza
economica, dal momento che nei Paesi non denutriti gran parte dell’alimentazione
č costituita da proteine.
Esistono
tre tipi di muscoli: muscolo striato scheletrico detto anche volontario; muscolo
striato cardiaco,
per fortuna involontario; muscolo liscio, anch’esso involontario e che al microscopio non presenta la
barratura trasversale caratteristica degli altri due, presente nel tubo
digerente, nella parete dei vasi sanguigni e in molti altri organi e apparati.
L’occhio
assonnato č spesso incluso tra le anomalie della cute e degli annessi cutanei.
Dato che si tratta di un’alterata funzione di un muscolo - salvo l’implicazione
di alterazioni neurologiche - mi pare doveroso inserire la mutazione se
- sleeping eye - in questo capitolo.
Non si tratta di sballottare i geni come fossero birilli.
Nel caso dell’occhio assonnato le notizie anatomopatologiche parlano solo di
alterata funzione del muscolo depressore della palpebra inferiore. Il
trasferimento di questa mutazione tra quelle di pertinenza neurologica sarŕ
possibile a patto di dati plausibili. Sta di fatto che i muscoli palpebrali
del pollo non appartengono neanche ai muscoli strettamente cutanei, o
pellicciai, come puň essere quello usato dal cavallo per scacciare le
mosche dal collo: il platěsma, dal greco platýs = piatto, ancora
presente anche nell’uomo nel quale mostra gradi variabili di sviluppo. Come
gli altri muscoli pellicciai, il platisma č praticamente privo di inserzioni ossee.
Invece i muscoli palpebrali del pollo si fissano, chi piů chi meno, a un osso. Nell’uomo non sono descritte inserzioni ossee per l’orbicolare
delle palpebre, che č quindi un muscolo essenzialmente cutaneo.
1.1. Occhio assonnato
|
se
- sleeping eye |
Autosomico
recessivo
Nel
pollo i movimenti delle palpebre sono assicurati dal muscolo orbicolare, dall’elevatore
della palpebra superiore e dal depressore di quella inferiore. Il muscolo
orbicolare si estende nello spessore delle palpebre inserendosi
perifericamente sull’osso lacrimale e sul mascellare, e con la sua contrazione
chiude le palpebre e ruota in avanti il bulbo oculare. Il muscolo elevatore
della palpebra superiore origina dall'osso frontale e si perde nella palpebra
omonima. Il muscolo depressore della palpebra inferiore si inserisce sul setto
interorbitale, in prossimitŕ del forame ottico, per arrivare sino al margine
libero della palpebra inferiore. La funzione di questi muscoli č insita nel
nome.
Bisogna tuttavia sottolineare che la palpebra superiore
svolge uno scarso ruolo nell'apertura della rima palpebrale, mentre gran
parte di questa funzione viene svolta dalla palpebra inferiore. Queste sono le
uniche notizie che ho potuto raccattare. Per analogia con l’uomo, vien da
pensare che anche nel pollo l’orbicolare serva a strizzare gli occhi. Vedrň
di sedermi tra i polli, per notare se anch’essi usano farsi l’occhiolino.
L’occhio
mezzo chiuso o occhio assonnato deriva da un’alterazione del
muscolo depressore della palpebra
inferiore che viene messo in movimento per aprire l’occhio. Di conseguenza
un’ipotonia di questo muscolo permetterebbe alla palpebra di risalire,
ottenendo cosě una parziale chiusura della rima palpebrale per un puro fatto
passivo. Insomma, la palpebra inferiore non serve a chiudere attivamente gli
occhi, ma ad aprirli. A mio parere la chiusura attiva č assicurata
dall’orbicolare. I libri di fisiologia del pollo non mancano, e hanno un
prezzo alquanto salato, ma morire se riesci ad arrivare a capo di un problema cosě
semplice! Chiedo scusa, ma io posso disquisire solo di medicina
umana. L’ultima parola sul pollo spetta ad altri.
I
pulcini portatori della mutazione se possono presentare delle rime
palpebrali
parzialmente o totalmente chiuse, ma quando vengono spaventati riescono ad
aprire gli occhi.
Una
situazione simile a quella determinata dal gene se č nota anche nell’uomo,
solo che č invertita, in quanto č la palpebra superiore a essere solo
parzialmente sollevata, per cui si parla di atteggiamento da astronomo:
il soggetto sta col capo reclinato all’indietro per avere una visione
migliore, e sembra stia sempre osservando il cielo con un telescopio. Questa
anomalia si riscontra spesso in membri della stessa famiglia, e
generalmente č dovuta a mancato sviluppo del muscolo elevatore della palpebra superiore
o ad aplasia dei nuclei di origine delle fibre nervose dell’oculomotore destinate
all’elevatore. Qualche volta nella ptosi congenita si riesce a mettere in
evidenza il fenomeno di Marcus Gunn: il soggetto riesce a sollevare la
palpebra ptosica tutte le volte che apre la bocca. Il fenomeno viene spiegato
ammettendo l’esistenza di anomale connessioni nervose.
Anche
nel pollo esiste un atteggiamento da astronomo, ma non da cause
palpebrali: il pulcino mantiene il capo
ruotato all’indietro e da un lato, col becco rivolto all’insů. Di quest’anomalia,
letale nel giro di pochi giorni, riparleremo a proposito della mutazione che
ne č responsabile, il gene loco, lo, nome preso dallo spagnolo nel
significato di pazzo.
Invece nell’occhio assonnato il pulcino mantiene
il capo reclinato in senso opposto rispetto a un astronomo e guarda sempre in avanti
verso terra. Quest’anomalia del pollo non scompare col progredire dell’etŕ
e anche se non č direttamente letale, č classificata come semiletale, in quanto
interferisce con l’alimentazione e la vita attiva.
1.2. Distrofia muscolare
|
am
- muscular dystrophy |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Furono
Asmundson & Julian (1956) a descrivere quest’affezione che non interessa
le fasi di sviluppo del muscolo, mentre ne compromette il trofismo. Gli
eterozigoti presentano masse muscolari piů voluminose del solito, con
alterazioni microscopiche identiche a quelle degli omozigoti.
I
pulcini presentano i primi sintomi dopo 2-3 settimane dalla nascita, e
consistono nell’incapacitŕ a mettersi in posizione normale quando vengono
rovesciati a pancia in su. Dal punto di vista anatomico č presente un aumento
di volume delle masse muscolari, cui segue un’atrofia nell’etŕ adulta,
con interessamento prima dei muscoli chiari e poi di quelli scuri.
Interferendo con la funzione dei sopracoracoidei, ne deriva un irrigidimento
delle articolazioni delle ali fino al loro blocco totale.
L’intimo
meccanismo di questa malattia genetica non č stato del tutto chiarito,
essendo in causa ora fattori insiti nella muscolatura, ora fattori di origine
nervosa che si esplicano a livello della giunzione neuromuscolare, cioč della sinapsi
neuromuscolare. Altre volte sembra responsabile un basso livello di
immunoglobuline circolanti.
Un’interpretazione
che potrebbe rispondere al vero č quella di Ralph Somes: si
tratterebbe di un’anomalia su base poligenica nella quale il gene am
esplica l’effetto maggiore,
dal momento che č stato possibile creare 4 differenti linee miodistrofiche
con caratteristiche ben distinte.
Il pollo affetto da questa patologia si č rivelato molto
utile negli studi biomedici, in quanto rappresenta un modello perfetto nella
prosecuzione della lotta alla stessa malattia ereditaria umana che attende un
chiarimento dalla ricerca scientifica.
1.3. Distrofia muscolare della Cornish
|
Distrofia
muscolare della Cornish |
Interazione
tra am e genotipo della Cornish
Fu Wagner (1970) a
studiare in una popolazione di Cornish una sindrome distrofica muscolare del
tutto simile alla precedente, nella quale tuttavia sono maggiormente
interessati i muscoli sopracoracoidei, pettorali e coracobrachiali, i quali
diventano giallastri e opachi. Pare plausibile l’ipotesi che sia in causa
ancora lo stesso gene am, e che le differenze siano legate al genotipo
della Cornish, capace di interagire con am. A differenza di quanto
accade nella distrofia muscolare classica, gli eterozigoti sono
indistinguibili dai soggetti normali.
1.4. Difetti del setto interventricolare
|
Difetti
del setto interventricolare |
Poligenico?
Recessivo con bassa penetranza?
Gruppo di associazione sconosciuto
Negli
animali i difetti del setto interventricolare sono le piů comuni anomalie
congenite a carico del cuore. Forma e dimensioni della comunicazione tra
ventricolo destro e sinistro sono variabili. Č stato dimostrato che nei
soggetti piů anziani molte comunicazioni si sono chiuse spontaneamente;
infatti solo i polli giovani dotati di anomalie piů severe presentano sintomi
clinici. Spesso un soggetto affetto da comunicazione interventricolare riesce
a riprodursi e a vivere una vita normale, anche se tollera meno la stagione
piů calda.
La
base genetica proposta si č spesso dimostrata abbastanza complicata e
potrebbe essere riassunta dicendo che questa condizione patologica č dovuta a
un’ereditarietŕ
di tipo recessivo
con bassa penetranza, oppure che sono coinvolti molteplici geni.
1.5. Cardiopatie differenti, alcune con possibili substrati genetici
So benissimo che qualcuno arriccerŕ il naso perché una parte di ciň che segue sembrerŕ ostrogoto autentico, ma non possiamo stilare un trattato di cardiologia propedeutica. Anche se succinto, richiederebbe troppe pagine. Mi metto a disposizione per qualsiasi chiarimento si rendesse necessario. Oltretutto la cardiologia č il mio pane quotidiano.
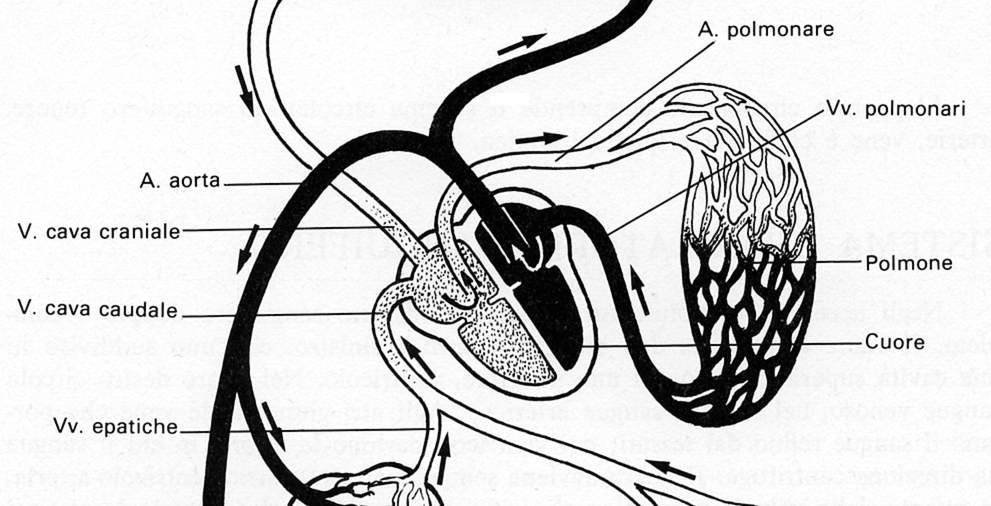
Cuore e sistema circolatorio degli Uccelli
Anche negli Uccelli si č evoluto un sistema circolatorio doppio e completo. Il cuore č contenuto nel sacco pericardico e comprende due distretti, destro e sinistro, ciascuno suddiviso in una cavitŕ superiore, atrio, e una inferiore, ventricolo, tra loro separate da un setto. Come negli uomini, nel cuore destro circola sangue venoso, nel sinistro sangue arterioso. Agli atri giungono le vene che portano il sangue refluo dai tessuti. Il sangue che giunge all'atrio destro deve ancora ossigenarsi, mentre quello che giunge all'atrio sinistro č andato a ossigenarsi nei polmoni. Dai ventricoli partono arterie il cui sangue ha direzione centrifuga. In condizioni normali il flusso avviene sempre in senso atrio-ventricolo-arteria, dal momento che le valvole presenti agli orifizi atrioventricolari e ventricoloarteriosi impediscono il reflusso del sangue.
Anche se un pollo abitualmente ha un solo cuore, tuttavia Siller (1967) si č preso la briga di raccogliere dati che potrebbero quasi farci inorridire: in un singolo soggetto Littré (1709) ne ha trovati 2, Mostafa (1952) ne ha contati 4 e Verocay (1905) nientemeno che 7. Larcher (1874) si č imbattuto in casi in cui le sezioni destre e sinistre del cuore avevano divorziato, cioč erano del tutto separate le une dalle altre. Quindi: addio setti interatriale e interventricolare.
La valvola interposta fra atrio e ventricolo sinistro č del tutto simile a quella dei mammiferi, solo che anziché di due lembi č composta da tre lembi per cui č una valvola tricuspide e non bicuspide o mitrale – a mitria rovesciata – come negli esseri umani.
La valvola atrioventricolare destra, che nei mammiferi č classicamente una tricuspide, negli uccelli č invece costituita da uno spesso lembo di tessuto muscolare foggiato a mezzaluna.
|
|
|
|
Cuore
di pollo – lato dorsale |
Cuore
di pollo – lato ventrale o sternale |
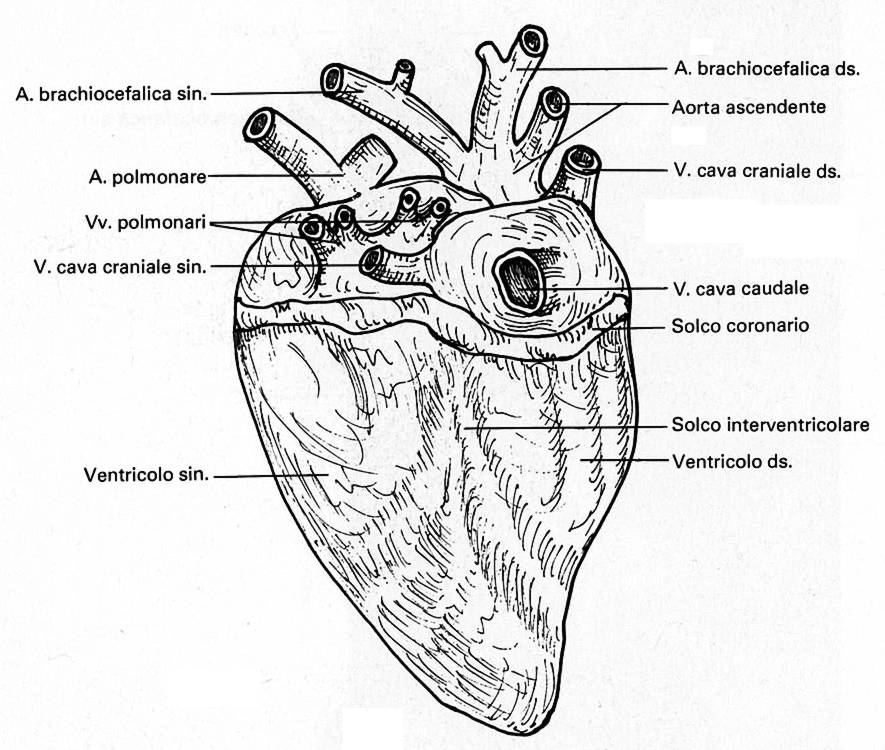
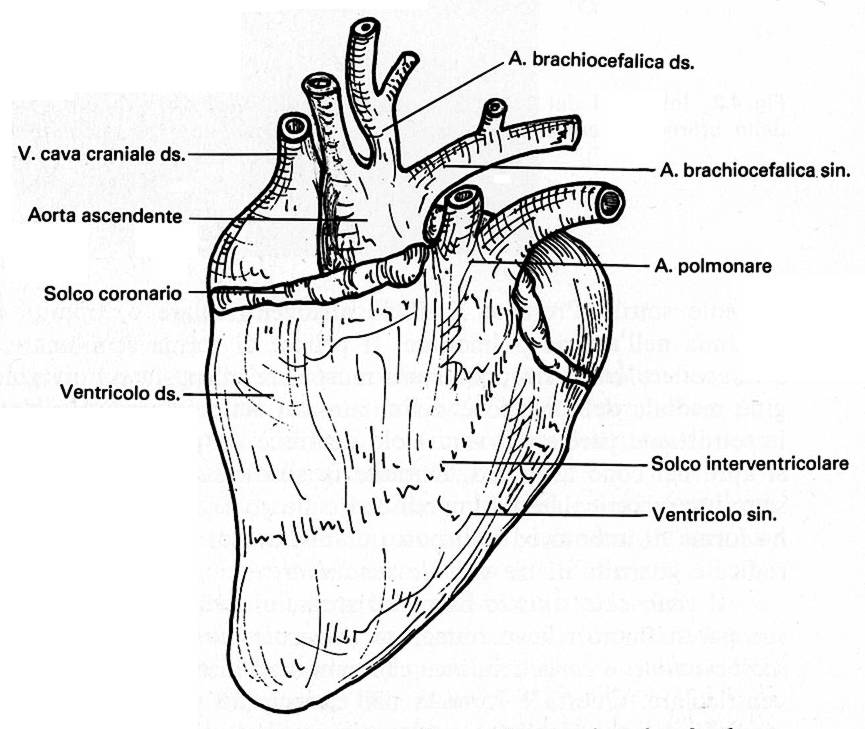
Č d'obbligo un breve cenno all'apparato di conduzione dell'eccitamento cardiaco. Esso č costituito, come nei mammiferi, dal nodo senoatriale e dal nodo atrioventricolare che si continua nel fascio atrioventricolare (A-V) di His, il quale a breve distanza dall'apice del cuore si divide in due branche, destra e sinistra, destinate alle pareti dei ventricoli.
Adesso iniziano le particolaritŕ dell'apparato di conduzione del cuore degli uccelli. Intorno all'orifizio A-V destro esiste l'anello di Purkinje che prende origine dal nodo A-V e che penetra appunto nella valvola A-V destra. Inoltre, dal fascio comune A-V di His si stacca una terza branca, il fascio ricorrente, il quale si porta verso l'ostio dell'aorta dove forma un anello atrioventricolare, e questo fascio ricorrente, dirigendosi posteriormente, unisce l'anello di Purkinje alla radice dell'aorta. Se non bastasse, un fascio derivato dalla branca destra del fascio di His penetra nella muscolatura della valvola A-V destra.
La frequenza cardiaca del pollo non č tra le piů basse, ma neppure tra le piů elevate presenti in alcuni uccelli: essa oscilla fra 350 e 470 battiti al minuto, e in effetti sono valori elevati se paragonati alla frequenza del cuore di un uomo adulto che batte in media 72 volte al minuto. Il pollo si trova a circa metŕ strada fra oca e canarino:
|
oca |
200 |
|
piccione |
221 |
|
tacchino |
200-280 |
|
pollo |
350-470 |
|
quaglia |
500-600 |
|
canarino |
1000 |
da Bell D.J. & Freeman B.M. Physiology and Biochemistry of the Domestic Fowl vol. 2 (1971)
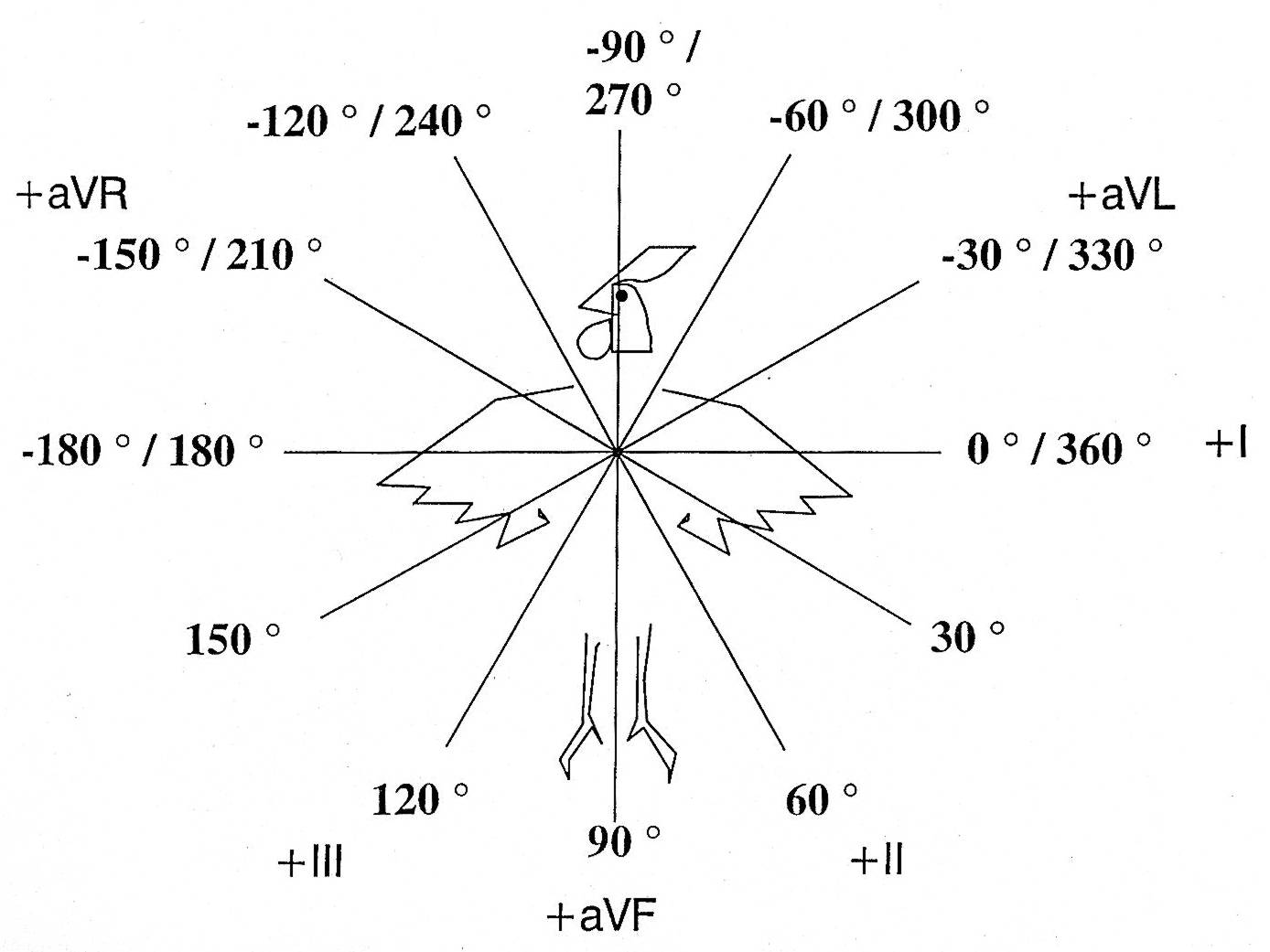
Direzione
assiale nel piano frontale
delle derivazioni bipolari I - II - III e unipolari aumentate aVR - aVL
- aVF
da Electrocardiographic diagnosis of cardiomyopathies in Aves di
Martinez, Jeffrey & Odom (1997)
Negli uccelli le cardiomiopatie possono essere studiate attraverso l'elettrocardiogramma – ECG -, come č stato accuratamente documentato da Martinez, Jeffrey & Odom in Electrocardiographic diagnosis of cardiomyopathies in Aves apparso nel 1997 in Poultry and Avian Biology Reviews. Un ECG di routine praticato a un uomo č costituito da 12 derivazioni: oltre alle 6 derivazioni unipolari toracice o precordiali, troviamo anche le derivazioni periferiche bipolari (I, II, III) e unipolari aumentate degli arti (aVR, aVL, aVF), mentre negli uccelli si registrano abitualmente solo queste ultime 6 derivazioni.
Nei volatili l'ECG puň servire allo studio di alcune affezioni cardiache: malattia del cuore tondo, sindrome da ipertensione polmonare o ascitica, sindrome della morte improvvisa, malattie infettive (colibacillosi, morbo di Newcastle), deficienze o eccessi nutrizionali.
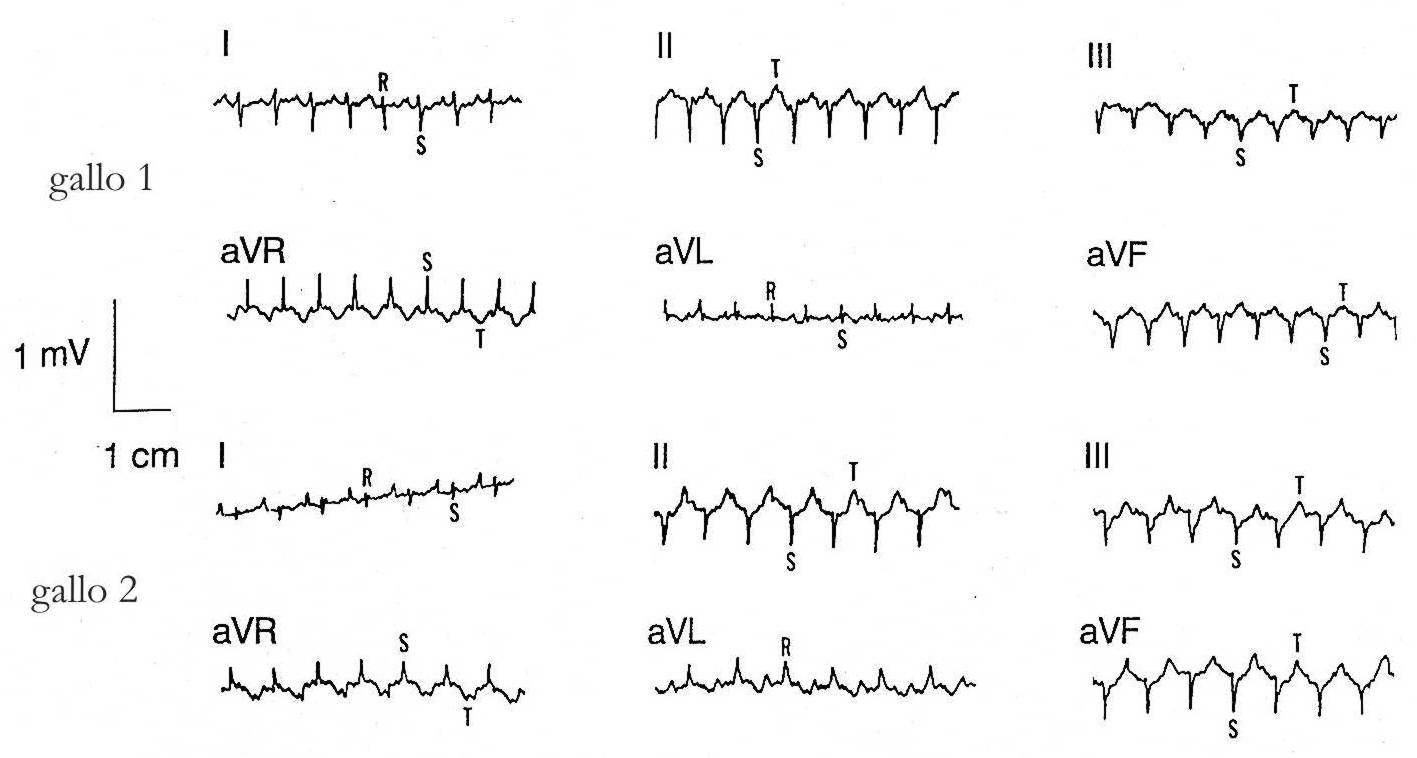
ECG
normale di due galli
derivazioni bipolari I - II - III e unipolari aumentate aVR - aVL - aVF
da Electrocardiographic diagnosis of cardiomyopathies in Aves di
Martinez, Jeffrey & Odom (1997)
Sindrome del cuore tondo
Anche se non interessa il pollo, vale la pena accennare alla sindrome del cuore tondo – round heart disease –, una miocardiopatia che colpisce prevalentemente i tacchini in etŕ compresa fra la 1Ş e la 10Ş settimana di vita. I soggetti presentano cianosi alle appendici carnose, penne arruffate, ali cadenti e ritardo dell'accrescimento. Il cuore si presenta arrotondato e notevolmente aumentato di volume per sfiancamento e assottigliamento del ventricolo destro. Il miocardio č pallido con i vasi coronarici dilatati, e talora nel pericardio si riscontra un versamento di liquido ambraceo. In addome puň essere presente un'ascite con materiale gelatinoso che riveste la superficie del fegato. La morte č in genere improvvisa. La causa sembra essere dovuta non a fattori genetici, bensě tossici, in quanto ricerche sperimentali hanno riprodotto il quadro morboso con l'impiego di furazolidone, un farmaco di norma usato nelle vaginiti e nelle enterocoliti umane. In questa patologia del tacchino l'ECG permette di seguire l'evolversi della malattia.
Sindrome da ipertensione polmonare
La sindrome da ipertensione polmonare - o sindrome ascitica - colpisce polli a rapida crescita come i broiler e consiste in un'insufficienza cardiaca congestizia che forse costituisce una minaccia economica temibile quanto l'influenza aviaria. La sindrome č caratterizzata da elevata pressione arteriosa polmonare, dilatazione e ipertrofia del ventricolo destro che sfocia in insufficienza cardiaca, con successiva dilatazione dell'atrio destro, stasi sanguigna e versamento di liquido in addome, cioč ascite. La causa risiederebbe in un'ipertensione idiopatica – cioč senza cause apparenti – a carico del letto vascolare polmonare. All'ECG si rileva un aumento di voltaggio dell'onda di depolarizzazione ventricolare e un aumento di ampiezza dell'onda S. Ulteriori particolari si possono dedurre dallo studio appena citato disponibile su richiesta. Anche se questa patologia č per definizione idiopatica – o essenziale – pare che l'ipossia del pulcino durante l'incubazione contribuisca alla sviluppo della sindrome.
Sindrome da morte improvvisa
La sindrome da morte improvvisa – sudden death syndrome, flip-over syndrome cioč sindrome del rovesciarsi – colpisce anch'essa i polli da carne in buone condizioni di nutrizione, i quali muoiono improvvisamente. Vengono trovati nella lettiera a pancia in su, quindi appoggiati sul dorso, con le zampe rivolte in alto e il collo esteso. All'autopsia non si riscontrano alterazioni degne di nota, se si eccettua per un certo numero di casi la presenza di congestione e di edema polmonare. Il tratto gastrointestinale appare costantemente pieno, il che sta a indicare la rapiditŕ con cui avviene la morte. Neppure a livello cardiaco esistono lesioni microscopicamente dimostrabili, nonostante oggi si propenda a classificare questa sindrome come una cardiomiopatia. Essa in genere colpisce galletti di 2-3 settimane d'etŕ, ma la sua eziologia non č stata ancora chiarita, anche se forse riconosce una base metabolica, dove entrano in gioco diversi fattori: genetici, nutrizionali (carenza di vitamine idrosolubili) e ambientali (eccesso di rumori e di luce). L'aggiunta alla dieta di biotina – vitamina H, un trasportatore di CO2 – riduce la mortalitŕ. Mentre stanno morendo, i galletti presentano all'ECG una fibrillazione ventricolare forse scatenata da un'aritmia ventricolare, aritmia che puň essere documentata come accade negli esseri umani. Per cui il poter rilevare con un ECG la presenza di aritmie ventricolari capaci di condurre alla fibrillazione ventricolare puň permettere di identificare i soggetti a rischio di morte improvvisa e quindi di effettuare una selezione per riproduttori non a rischio. Il che mi pare un ottimo motivo economico che giustifica appieno la registrazione dell'ECG nei polli. Infatti qualcuno potrebbe protestare di fronte a un pollo che si sdraia bellamente sul lettino per farsi fare un ECG senza alcun intoppo burocratico e magari per motivi in apparenza futili o accademici, mentre un essere umano č spesso costretto a chiedere l'elemosina se vuol prenotare un ECG senza attese quaresimali!
Tralascio altri dettagli rilevabili e documentabili all'ECG sia in corso di infezioni che di deficit vitaminico-minerali, in quanto non strettamente pertinenti dal punto di vista genetico.
Combattenti della Sonda sconfitti dalla morte improvvisa
Prima di chiudere questo rapido excursus cardiologico non posso non rendervi partecipi di un'esperienza personale che in parte concorda e in parte discorda con ciň che per ora ci č dato sapere circa l'eziopatogenesi della sindrome da morte improvvisa. La concordanza sta nel fatto che due galli combattenti della Sonda, padre e figlio, mi sono morti improvvisamente.
Il padre č morto davanti ai miei occhi e mi ha lasciato di stucco. Ormai da qualche tempo gli era venuta la mania di pretendere che fossi suo partner quando ci incontravamo in pollaio tutti i giorni. Il figlio invece mi ha sempre ignorato. Agli inizi il padre cominciň ad aggredirmi con le zampe sforbiciandole sempre piů vicine alle mie gambe, finché non lo catturavo e lo lanciavo fuori in frutteto. Motivi di rispetto per il suo patrimonio genetico esigevano tuttavia di assecondarlo, per cui decisi di portare in pollaio un paio di guanti piuttosto robusti. Dopo aver raccolto le uova e messo il cibo nelle mangiatoie, afferravo i guanti dal lato opposto alle dita e glieli sventolavo davanti per un po' di tempo, un tempo che per lui non era mai troppo, e il marpione si trastullava ad aggredirli con colpi di speroni. Un giorno uscendo dal pollaio lui mi segue e mi accompagna verso la parte bassa del frutteto, esigendo a ogni pie' sospinto che lo faccia divertire. All'ennesima scaramuccia coi guanti ecco che alza come al solito le zampe da terra, ma subito crolla riverso, flip-over. Midriasi e assenza di riflesso corneale mi hanno trattenuto dal praticargli un massaggio cardiaco, magari accompagnato da una respirazione becco a bocca. L'ho raccolto e l'ho sepolto con grande rimpianto e disappunto.


Coppia
di Combattente della Sonda identica alla mia
che non ebbi mai modo di fotografare
da Handbuch der Hünerrassen (1996) - foto di Josef Wolters
Anche il figlio č morto improvvisamente, e non era un giovincello, cosě come non lo era il padre, e ambedue non hanno mai avuto eccesso di luce o di rumori. Tutt'altro. E di vitamine ne avevano a iosa, tutte quelle che madre natura sa elargire. Era una luminosa e gelida giornata di gennaio. Verso l'una del pomeriggio vedo dalla finestra che figlio Sonda se ne sta coricato al sole, quando i polli si dedicano a una pausa prima di chiudere la giornata con qualche battibecco o con qualche scopata. Il classico momento in cui galli e galline si radunano tranquillamente come farebbe una grande pacifica famiglia. Lui era discosto dal gruppo, sembrava si godesse la pennichella in solitudine e che prendesse il sole, oppure – mi dissi memore del padre – se ne sta sdraiato perché č morto. Avrei verificato piů tardi, in quanto il freddo era pungente e non avevo voglia di imbottirmi di giacche e cappotti. Se č morto č morto. Amen. Infatti, in occasione del consueto giro pomeridiano, lo trovo cadavere. Morte improvvisa come il padre. E seppellii anche lui con altrettanto disappunto.
Se il padre č morto flip-over, č perché stava aggredendo i guanti. Il figlio non era flip-over, ma disteso a pancia in giů e spaparacchiato come un bagnante sulla spiaggia. Inoltre, ambedue non erano pivelli, e ambedue non erano incorsi in quei fattori nutrizionali e ambientali cui sono soggetti i broiler perennemente carcerati.
Ma padre e figlio avevano un qualcosa in comune: il corredo genetico, forse la causa del tutto.
2. Testa
2.1. Ernia cerebrale e bozza cranica
|
Ernia
cerebrale e bozza cranica |
Dovuta
a Cr e a geni modificatori
Un’ernia
cerebrale, alloggiata in una bozza della calotta cranica, č spesso
associata al ciuffo, specialmente in quei polli dotati di un ciuffo grande.
In The variation of animals and plants under domestication (1868) Charles Darwin ci fornisce un ampio resoconto sull’ernia
cerebrale associata a bozza cranica dei polli ciuffati. Ai suoi tempi tutte
le varietŕ di Polish possedevano una grande protuberanza cranica sviluppata
in egual misura in ambo i sessi, mentre in Germania in tempi anteriori solo
il cranio della gallina era dotato di protuberanza, come aveva affermato Johann
Friedrich
Blumenbach![]() nel
1805 pubblicando i suoi studi anatomici sulle anormalitŕ degli animali
domestici. Anche Johann
Matthäus
Bechstein
nel
1805 pubblicando i suoi studi anatomici sulle anormalitŕ degli animali
domestici. Anche Johann
Matthäus
Bechstein![]() nel 1793 aveva fatto la stessa osservazione di Blumenbach e non si
era limitato a riferire sugli effetti del ciuffo solamente nel pollo, ma
anche nell’oca, nell’anatra e nel canarino. Vista l’autorevolezza e la
serietŕ degli studiosi tedeschi, Darwin conclude che senza dubbio in
Germania la bozza cranica della Polish un tempo era confinata al sesso
femminile e che successivamente si č trasferita anche al gallo, cosě da
costituire un tratto comune ai due sessi.
nel 1793 aveva fatto la stessa osservazione di Blumenbach e non si
era limitato a riferire sugli effetti del ciuffo solamente nel pollo, ma
anche nell’oca, nell’anatra e nel canarino. Vista l’autorevolezza e la
serietŕ degli studiosi tedeschi, Darwin conclude che senza dubbio in
Germania la bozza cranica della Polish un tempo era confinata al sesso
femminile e che successivamente si č trasferita anche al gallo, cosě da
costituire un tratto comune ai due sessi.
Fisher
(1935) fornisce
un’interpretazione genetica di quanto affermato da Bechstein e Blumenbach
e che viene accettato da Darwin: durante la domesticazione l’ernia
č stata soppressa nella varietŕ giapponese della Silky da un
fattore o da fattori ereditari capaci di sopprimere l’ernia piů facilmente
nel maschio rispetto alla femmina. Sempre secondo Fisher sembra che in
passato altri polli ciuffati, come la Polish, siano stati affetti da una
tale soppressione genetica, che venne poi eliminata quando gli allevatori
amatoriali si prefissero il traguardo di un bel cranio a cupola.
Darwin
fornisce un’altra notizia storica sullo studio del cranio della Polish,
affermando che: “The anatomical peculiarities of the skull of the Polish
breed were noticed by P. Borelli in 1656.” (pag.
260
della seconda edizione revisionata dallo stesso Darwin). Č una svista onomastica piů o meno latineggiante
di Darwin, in quanto non si tratta di un Borelli italiano, bensě del
francese Pierre Borel![]() (circa 1620-1671) latinizzato in Petrus Borellus, protestante, consigliere oltre
che medico alla Corte francese. In base al caleidoscopico contenuto delle
sue opere doveva possedere una mente assai versatile che spaziava dalla
linguistica alla microscopia. Infatti il primo lavoro medico basato
sull’uso del microscopio č il suo Historiarum et observationum
medicophysicarum centuriae
IV ed č nella seconda centuria -
raccolta di cento cose dello stesso genere – risalente piů o meno al 1653
che Pierre Borel accenna alla foggia del cranio del pollo ciuffato. Infatti,
oltre a un gatto con due teste, egli conservava anche il cranio di una
gallina che si presentava come composto da due crani uno aderente
all’altro, uno dei quali era informe: “Felem bicipitem etiam servo, ut et
cranium gallinae, cui quasi aliud cranium informe adhaeret.”
(centuria II – observatio LVII – Monstrum biceps,
& testudineus tumor mirus) Questa
preziosa identificazione e citazione di Pierre Borel č dovuta alle ricerche
sui polli ciuffati di Elisabetta Maresio (2005) ed č tratta da uno studio del 1839
sui polli ciuffati di E. Hagenbach
(circa 1620-1671) latinizzato in Petrus Borellus, protestante, consigliere oltre
che medico alla Corte francese. In base al caleidoscopico contenuto delle
sue opere doveva possedere una mente assai versatile che spaziava dalla
linguistica alla microscopia. Infatti il primo lavoro medico basato
sull’uso del microscopio č il suo Historiarum et observationum
medicophysicarum centuriae
IV ed č nella seconda centuria -
raccolta di cento cose dello stesso genere – risalente piů o meno al 1653
che Pierre Borel accenna alla foggia del cranio del pollo ciuffato. Infatti,
oltre a un gatto con due teste, egli conservava anche il cranio di una
gallina che si presentava come composto da due crani uno aderente
all’altro, uno dei quali era informe: “Felem bicipitem etiam servo, ut et
cranium gallinae, cui quasi aliud cranium informe adhaeret.”
(centuria II – observatio LVII – Monstrum biceps,
& testudineus tumor mirus) Questa
preziosa identificazione e citazione di Pierre Borel č dovuta alle ricerche
sui polli ciuffati di Elisabetta Maresio (2005) ed č tratta da uno studio del 1839
sui polli ciuffati di E. Hagenbach![]() .
.

Pierre Borel - Historiarum et observationum
medicophysicarum Centuriae IV
Centuria II – Observatio LVII

In tempi piů recenti fu Tegetmeier (1856) a descrivere la situazione anatomica del cranio nella Polish, nella Silky e nella Houdan. Quindi fu appunto la volta di Darwin (1868) che da ottimo naturalista esaminň ben 14 crani appartenenti a Polish e ad altri polli ciuffati. Ecco quali furono i reperti a carico dei 9 polli appartenenti a varietŕ di Polish inglese: in sezioni longitudinali, condotte contemporaneamente in un cranio di Cochin delle stesse dimensioni allo scopo di avere un confronto, la protuberanza emisferica delle ossa frontali occupava nelle due razze la stessa posizione, ma nella Polish aveva dimensioni alquanto diverse. Inoltre, sempre in un esemplare di Polish, l’osso era estremamente sottile; il grado di ossificazione della protuberanza variava parecchio, in quanto porzioni piů o meno ampie erano sostituite da una membrana; in un soggetto esisteva solamente un singolo foro aperto, mentre per lo piů esistevano spazi aperti di morfologia variabile, tanto da dare l’aspetto di un osso reticolato; per lo piů esisteva un nastro di osso arcuato a disposizione longitudinale e mediana, ma in un caso, pulito il cranio, questo si mostrň come una fossetta vista dall’alto, in quanto mancava qualsiasi traccia di osso.
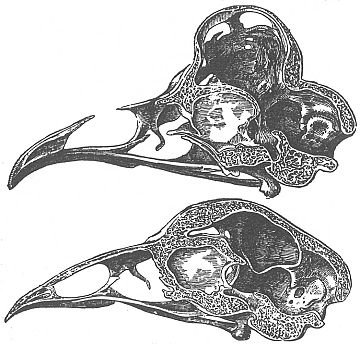
Fig.
X. 1 - Darwin: cranio di gallo Polish a confronto con quello di un gallo
Cochin.
Il
cervello era modificato in modo conseguente e andava a occupare la cavitŕ
anormale. Da notare inoltre che nella Cochin e nei polli abituali un robusto
setto osseo separava la cavitŕ cranica anteriore da quella media, mentre
nel cranio di Polish raffigurato questo setto era assente e anche la
cavitŕ media, che nella Cochin era allungata, apparve a Darwin di forma
circolare, in quanto si trattava di una sezione longitudinale. Una domanda
che Darwin si fece, e che altri gli posero, č se questa situazione
anatomica del cervello potesse influenzare le capacitŕ psichiche della
Polish, in quanto spesso si dice che questa razza č stupida e alquanto
idiota. Tegetmeier era di questo avviso, mentre Darwin si limitň a
descrivere il comportamento di alcuni suoi soggetti, che invero non era del
tutto normale.
Come riferisce Hagenbach che abbiamo testé citato
parlando di Pierre Borel, giŕ nel 1767 il naturalista tedesco Peter Simon Pallas (Berlino 1741-1811)![]() nel primo volume dei suoi Spicilegia zoologica affermava che
si trattava di polli stupidi e inetti: “Notabile est, et ad confirmandam
meam opinionem facit, quod Gallinae omnes hacce cranii praeternaturali
conformatione vitiatae stupidae quasi et ineptae esse videantur nec
longaevae sint, omnesque per domesticam vitam summopere corruptae
constitutionis notas prae se ferre soleant.” A Pallas faceva eco qualche
decennio piů tardi un altro naturalista tedesco, Johann Friedrich Blumenbach
nel primo volume dei suoi Spicilegia zoologica affermava che
si trattava di polli stupidi e inetti: “Notabile est, et ad confirmandam
meam opinionem facit, quod Gallinae omnes hacce cranii praeternaturali
conformatione vitiatae stupidae quasi et ineptae esse videantur nec
longaevae sint, omnesque per domesticam vitam summopere corruptae
constitutionis notas prae se ferre soleant.” A Pallas faceva eco qualche
decennio piů tardi un altro naturalista tedesco, Johann Friedrich Blumenbach![]() (Gotha 1752-Gottinga 1840), uno dei fondatori dell'antropologia come scienza oggettiva, il quale
intorno al
1812 affermava che questi polli erano un po’ stupidotti e dementi: “Notus
vero ceteroquin effectus, quem gravis haec cerebri deformatio in animae
facultates harum gallinarum exserit, utpote quas tantum non semper etsi
vario gradu stupidas et quasi dementes esse constat.”
(Gotha 1752-Gottinga 1840), uno dei fondatori dell'antropologia come scienza oggettiva, il quale
intorno al
1812 affermava che questi polli erano un po’ stupidotti e dementi: “Notus
vero ceteroquin effectus, quem gravis haec cerebri deformatio in animae
facultates harum gallinarum exserit, utpote quas tantum non semper etsi
vario gradu stupidas et quasi dementes esse constat.”
Esiste
anche un altro problema anatomico correlato alla bozza del cranio. Nella
maggioranza dei polli il processo postero-laterale del frontale e il
processo dello squamoso sono ossificati alle loro estremitŕ, ma l’unione
dei due ossi non č costante in tutte le razze. In 11 dei 14 crani
esaminati, questi processi erano nettamente distinti, e, quando questo
accade, invece di essere inclinati anteriormente, scendevano ad angolo retto
verso la mandibola, per cui l’asse maggiore della cavitŕ ossea dell’orecchio
risulta piů perpendicolare.
Esiste
poi il problema delle ossa nasali. In condizioni normali ogni osso nasale si
spinge anteriormente, con due estesi processi di uguale lunghezza, tanto da
formare una forca. Orbene, nella Polish, il processo mediale era accorciato
e rigirato all’insů in grado variabile. A questo si deve aggiungere che
anche la branca ascendente del premascellare, invece di spingersi tra le
branche mediali delle due ossa nasali, era molto accorciata e terminava in
una punta rivolta in alto, senza poter raggiungere l’etmoide e il
frontale. Ne consegue che le coane sono piů grandi e a forma di mezzaluna.
Non contento, Darwin esaminň il cranio di due Sultano e trovň che la protuberanza della femmina era maggiore di quella del maschio. In ambedue i soggetti la branca ascendente del premascellare era molto breve e la parte mediale del processo dell’osso nasale bilaterale era fusa con l’altra e ossificata.
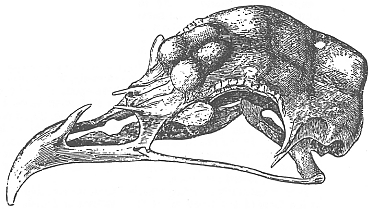
Fig.
X. 2 - Darwin: cranio di Pollo Cornuto.
Tegetmeier
inviň a Darwin il cranio di un Pollo Cornuto, cioč con cresta a cornetti.
Darwin lo classificň come una sottorazza della Polish e gli diede il nome
di Horned fowl: era dotato di un piccolo ciuffo nonché di una cresta
costituita da due grandi punti, ciascuno dei quali era supportato da due
protuberanze ossee. Il cranio di questo Pollo Cornuto presentava delle
caratteristiche simili, sotto molti profili, a quelle della Polish, ma non
aveva la grande protuberanza del frontale. Al suo posto si potevano appunto
notare quattro piccole protuberanze, due per lato, disposte piů
anteriormente, al di sopra delle ossa lacrimali. Queste nocche, nelle quali
il cervello non penetrava, erano tra loro separate da un profondo solco mediano
bucherellato da pochi fori minuti. La situazione dello scheletro delle coane
non era differente da quello della Polish. Senza dubbio alcuno, queste
protuberanze facevano da supporto alla cresta conformata a cornetti.
Il
ciuffo si accompagna a situazioni anatomiche craniche che variano con le sue
dimensioni, tant’č che, secondo Bechstein, in presenza di un ciuffo
modesto si poteva osservare anche solo un’escrescenza carnosa senza
alcuna protuberanza dell’osso cranico. Darwin aggiunge che ha potuto
osservare una tale escrescenza, carnosa o fibrosa, nelle anatre ciuffate,
nelle quali esisteva solamente un cranio leggermente globoso.
Cosě,
attraverso la selezione, l’uomo ha via via accentuato la protuberanza
cranica con tutte le anomalie connesse a carico del naso e dell’asse della
cavitŕ auricolare. Da non dimenticare che anche la morfologia del cervello
ha subito modificazioni secondarie.
Per chi fosse interessato ad approfondire cosa
accade al cervello di un pollo ciuffato possiamo consigliare tre lavori
scientifici estremamente specialistici ma non per questo da mettere nel
dimenticatoio, in quanto la scienza ha l’obbligo di scendere nei minimi
dettagli servendosi della tecnologia del momento. Il primo lavoro č quello
di E. Hagenbach giŕ citato e
che risale al
1839![]() .
Il secondo
č del 1959 e precisamente č lo studio assai approfondito e documentato di
Horst Requate che riguarda altri volatili e non solo il pollo ciuffato: Federhauben
bei Vögeln. Eine genetische und
entwicklungsphysiologische Studie zum Problem der Parallelbildungen (Z. Wiss.
Zool., 162: 191-313). Il terzo č piů recente e risale al 1998: Allometric Comparison of
the Brain and Brain Structures in the White Crested Polish Chicken with
Uncrested Domestic Chicken Breeds di Heiko D. Frahm, Gerd Rehkämper del
C. und O. Vogt Institute of Brain Research, University of Düsseldorf,
Germany (in Brain, Behavior and Evolution 1998; 52:292-307). Frahm & Rehkämper giungono a questa
conclusione: certe strutture cerebrali della Polish ciuffo bianco non sono
significativamente piů grandi rispetto a quelle di polli non ciuffati,
mentre in questa razza ciuffata il tratto ottico, il diencefalo, il
telencefalo, l’iperstriato accessorio, l’iperstriato dorsale e ventrale
nonché il neostriato hanno dimensioni significativamente aumentate
.
Il secondo
č del 1959 e precisamente č lo studio assai approfondito e documentato di
Horst Requate che riguarda altri volatili e non solo il pollo ciuffato: Federhauben
bei Vögeln. Eine genetische und
entwicklungsphysiologische Studie zum Problem der Parallelbildungen (Z. Wiss.
Zool., 162: 191-313). Il terzo č piů recente e risale al 1998: Allometric Comparison of
the Brain and Brain Structures in the White Crested Polish Chicken with
Uncrested Domestic Chicken Breeds di Heiko D. Frahm, Gerd Rehkämper del
C. und O. Vogt Institute of Brain Research, University of Düsseldorf,
Germany (in Brain, Behavior and Evolution 1998; 52:292-307). Frahm & Rehkämper giungono a questa
conclusione: certe strutture cerebrali della Polish ciuffo bianco non sono
significativamente piů grandi rispetto a quelle di polli non ciuffati,
mentre in questa razza ciuffata il tratto ottico, il diencefalo, il
telencefalo, l’iperstriato accessorio, l’iperstriato dorsale e ventrale
nonché il neostriato hanno dimensioni significativamente aumentate![]() .
Tanto basti per invogliare i curiosi ad approfondire l’argomento.
.
Tanto basti per invogliare i curiosi ad approfondire l’argomento.
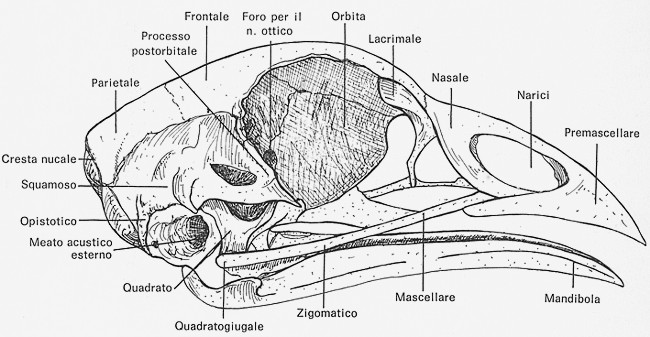
Fig.
X. 3 - Cranio di un pollo abituale visto lateralmente
Nel
1979
Don Brothwell ha
pubblicato un articolo molto interessante che sicuramente costituisce
una pietra miliare per la storia della genetica del ciuffo e dell’ernia
cerebrale: Roman
evidence of a crested form of domestic fowl, as indicated by a skull showing
associated cerebral hernia. Nel 1977 tra
i rifiuti del complesso di un tempio Romano Britannico situato a West Hill
di Uley nel Somerset (UK) vennero alla luce scheletri di animali, il 2% dei quali apparteneva al genere Gallus. Da una moneta si
puň desumere che il tempio potrebbe essere stato costruito nel secondo
quarto del IV secolo dC per essere poi demolito verso il 380 dC. Il cranio
descritto da Brothwell nel suo lavoro č identificato come WH77(162)CH ed č
l’unico tra i crani di Gallus ritrovati a Uley a presentare uno sviluppo
anormale. Si tratta di un cranio quasi completo al quale mancano sia l’area
nasale, premascellare, mascellare e zigomatica, nonché la mandibola. Esiste
qualche ulteriore danneggiamento nel punto in cui il frontale avrebbe dovuto
estendersi sopra l’etmoide oltre che sopra l’orbita sinistra;
esternamente, sia a destra che a sinistra del foramen magnum, cioč
del foro occipitale, l’aspetto presenta alcuni danni, ma visto da dietro
questo cranio si presenta del tutto simile ad altri crani di pollo ritrovati a Uley e
in altri siti. Visto di lato e dall’alto, questo cranio presenta una
notevole espansione globosa del frontale che si estende dall’area nasale
fin verso la regione parietale. Esistono almeno due grandi fori visibili
frontalmente e che risalgono a prima della morte del pollo: uno di questi -
il destro - č osservabile anche lateralmente ed č di gran lunga
maggiore essendo alto 11,3 mm e largo 7 mm, mentre il sinistro misura 5x3,9
mm.
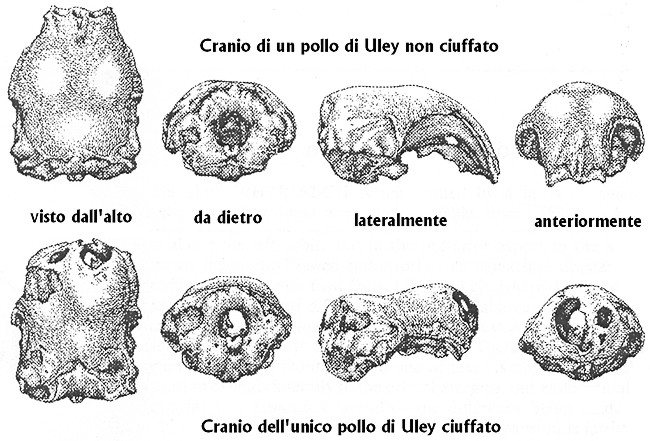
Fig. X. 4 - Crani di polli
romani di Uley del IV secolo dC (da Brothwell, modificata). A proposito del cranio
dell’unico pollo ciuffato di Uley possiamo fare altre due considerazioni:
visto da dietro, il foro occipitale permette di scorgere l’ampia
fenestratura destra a carico del frontale; visto di fronte, tale
fenestratura lascia intravedere il foro occipitale. Sembra quasi che i
reperti di Brothwell a carico del cranio del pollo ciuffato vogliano
illustrare i reperti di Darwin del 1868 che citiamo nuovamente: “Inoltre,
sempre in un esemplare di Polish, l’osso era estremamente sottile; il
grado di ossificazione della protuberanza variava parecchio, in quanto
porzioni piů o meno ampie erano sostituite da una membrana; in un soggetto
esisteva solamente un singolo foro aperto, mentre per lo piů esistevano
spazi aperti di morfologia variabile, tanto da dare l’aspetto di un osso
reticolato; per lo piů esisteva un nastro di osso arcuato a disposizione
longitudinale e mediana, ma in un caso, pulito il cranio, questo si mostrň
come una fossetta vista dall’alto, in quanto mancava qualsiasi traccia di
osso.”
Brothwell conclude che non esiste alcun dubbio sul fatto che tale cranio mostri una spiccata espansione del frontale come risultato di un’ernia cerebrale. Nonostante in base ai dati esposti - prosegue Brothwell - sia incorretto proporre che qualcuna delle moderne varietŕ di polli ciuffati fosse posseduta dai Romani, si puň tuttavia affermare che si tratta della prima evidenza a supporto del fatto che oltre 1500 anni fa esisteva un pollo ciuffato. Poi, lo stabilire se si trattasse di una varietŕ ben stabilizzata dal punto di vista genetico e fenotipico oppure se si trattasse di un’occasionale mutazione individuale, č un problema che potrŕ essere risolto solo attraverso lo studio di ulteriori reperti romani di pollo.
Fu Krautwald (1910)
a dimostrare che la genesi dell’ernia cerebrale consiste in un accumulo di liquido cefalorachidiano nel terzo e quarto
ventricolo,
capace di determinare una protrusione degli emisferi cerebrali, cui si
sovrappongono successivamente le ossa frontali.
Non si conoscono le relazioni esistenti tra bozza e ciuffo. Tuttavia una relazione č certa: quanto piů il ciuffo č grande tanto piů l’anomalia cranica č estesa. Nel paragrafo seguente troveremo l’interpretazione genetica del ciuffo di Warren & Hutt. E, come vedremo a proposito del locus D responsabile della cresta doppia, pare che le narici piů ampie e cavernose si associno a uno stato di omozigosi Dv/Dv.
2.2. Ciuffo e ernia cerebrale
|
Cr
- crest |
Autosomico
incompletamente dominante
che agisce insieme a geni modificatori indipendenti
per ciascuno dei due caratteri
Gruppo di associazione II - cromosoma 2
Bisogna
ricordare che alcune razze dotate di fiocco
o di ciuffo
a elmo,
come Barbuta di Watermaal ![]() -
- ![]() , Brabantea
[1]
, certe
varietŕ di Combattente Inglese Antico nano, Ciuffata di Appenzell
, Brabantea
[1]
, certe
varietŕ di Combattente Inglese Antico nano, Ciuffata di Appenzell ![]() , non
mostrano un’ernia cerebrale, che sarebbe motivo di squalifica.
, non
mostrano un’ernia cerebrale, che sarebbe motivo di squalifica.
Le basi genetiche dell’ernia cerebrale non sono state ancora del tutto chiarite. L’ernia č sempre stata strettamente associata alla presenza di un ciuffo e in passato le due manifestazioni furono attribuite a due distinti geni recessivi, ma Warren e Hutt (1936) - sulla scia degli esperimenti di Fisher (1934, 1935) - dimostrarono che ambedue i tratti sono dovuti a un singolo gene Cr, e che ciascuno di questi tratti viene interessato in modo indipendente da geni modificatori che conducono a fenotipi molto variabili. I due ricercatori dimostrarono pure che il locus Cr č associato al locus del bianco dominante I, distante 12,5 unitŕ crossover. I soggetti eterozigoti per Cr generalmente non presentano ernia cerebrale. I ciuffati con cresta semplice e i ciuffati con cresta a rosa non presentano ernia cerebrale.
Una trattazione esauriente del ciuffo si trova nel capitolo che riguarda le mutazioni attive sul piumaggio; per un'analisi storica sulla probabile epoca di comparsa di questa mutazione si veda l'ultimo capitolo del III volume.
2.3. Tuberositŕ del cranio
|
KfH - kuif |
Autosomico
dominante
Gruppo di associazione II - cromosoma 2
Requate
(1960),
studiando la Polish nana ciuffo bianco ![]() -
- ![]() , giunse alla conclusione che la
tuberositŕ del cranio non č dovuta a Cr, ma a un gene separato
dominante KfH
posto molto vicino al precedente, dal quale dista solo 3,5 unitŕ.
, giunse alla conclusione che la
tuberositŕ del cranio non č dovuta a Cr, ma a un gene separato
dominante KfH
posto molto vicino al precedente, dal quale dista solo 3,5 unitŕ.
2.4. Interazione fra ernia cerebrale e tipo di cresta
Un
aspetto dello sviluppo dell’ernia cerebrale, che non pare indagato in
nessuno degli studi pubblicati, č la possibile
interazione fra il tipo di cresta e l’espressione dell’ernia
cerebrale. Le
ernie associate ad ampio ciuffo vanno di pari passo con creste molto piccole,
duplici, a forma di lettera V. Ciň potrebbe
spiegarsi col fatto che l’ernia puň esprimersi quando la sommitŕ del capo
č piů o meno priva della massa carnosa dovuta alla cresta. Come diremo a
proposito della cresta doppia dovuta al locus D, pare possibile
l'associazione fra i geni Dv e Dc e
un'ernia cerebrale.
2.5. Becco incrociato
|
Becco
incrociato |
Ereditarietŕ
recessiva multifattoriale con penetranza ridotta
Gruppo di associazione sconosciuto
Parecchie
razze mostrano questo difetto, talora associato a microftalmia oppure ad
anoftalmia monolaterale, ma queste anomalie oculari non sono dovute a cause
genetiche, bensě a uno sviluppo anomalo del bulbo, secondario a sua volta
alla malformazione ossea. I due tipi di becco a croce, semplice o associato a
difetti dell’occhio, vengono spesso influenzati
dalle condizioni in cui si svolge l’incubazione. Per lo piů il pulcino nasce con un becco normale che comincia a
incrociarsi verso 1-2 mesi d’etŕ, raggiungendo l’espressione massima a
3-4 mesi.
L’anomalia
puň interessare sia la parte superiore che inferiore del becco, ma la
parte piů spesso colpita č quella superiore,
che puň presentarsi curvata lateralmente, per lo piů a destra, puň esserne
interessata solo la punta oppure tutta quanta la sua estensione. Anche il
cranio č deformato e asimmetrico, ed č ritenuto la causa delle anomalie del
becco. Un’osservazione
attenta puň mettere in evidenza anche un’asimmetria a carico dello sviluppo
e della struttura della lingua, che presenta una punta corneificata e una
superficie ricoperta da epitelio eccessivamente cheratinizzato.
Le
ricerche volte a dimostrare la base genetica di quest’anomalia non hanno
fornito dati univoci. Alcuni studiosi, tra cui Hutt, ritengono si tratti di un
gene recessivo; dello stesso avviso č Landauer, colui che nel 1938 ha
compiuto gli studi piů approfonditi, dimostrando che, pur ricorrendo a una stretta consanguineitŕ, non
sempre si riusciva a ottenere la deformitŕ. Landauer concluse per un’ereditarietŕ di tipo
recessivo, ma complessa. D’altro canto, ottenne il becco a croce iniettando
etilcarbamato.
Un’altra situazione studiata da Landauer, e riscontrata
solo nella Leghorn, č quella in cui i pulcini presentano il becco a croce fin
dalla nascita, ma nei quali il difetto si corregge coll’avanzare dell’etŕ.
L’incrocio tra questo tipo e il tipo precedentemente descritto, condusse a
una progenie normale. Per l’anomalia presente alla schiusa č stata proposta
una base recessiva multifattoriale con penetranza ridotta.
2.6. Brevitŕ della mandibola
|
sm - short
mandible |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Si tratta di una condizione semiletale in cui la mandibola
č ridotta di circa la metŕ rispetto alla lunghezza del becco superiore, per
cui la bocca rimane aperta in modo permanente e i pulcini si nutrono con
difficoltŕ. Quelli che riescono a sopravvivere, in etŕ adulta presentano una
lunghezza della mandibola che si avvicina a quella di 5-6 mesi d’etŕ.
Siccome il becco superiore si incurva verso il basso, č necessario tagliarlo
frequentemente. L’espressivitŕ di sm č variabile, in quanto da un
incrocio tra omozigoti sono nati 2 soggetti normali e 37 menomati.
2.7. Brevitŕ del becco superiore
|
su
- short upper beak |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Questa
malformazione colpisce, oltre al becco superiore, anche gli arti, piů spesso
le ossa lunghe delle ali rispetto a quelle delle gambe, pur non esistendo una
correlazione tra entitŕ di accorciamento del becco e brevitŕ delle ossa
lunghe delle appendici.
Il
becco varia da 1 mm di lunghezza sino ad essere completo. La schiusa č
percentualmente bassa, aggirandosi intorno al 13%, e la mortalitŕ postnatale
č legata a problemi di nutrizione. Tra coloro che riescono a raggiungere la
maturitŕ, alcuni continuano a presentare un becco breve, mentre altri
sviluppano un becco di normali dimensioni. Altri ancora hanno successivamente
sviluppato un becco a croce, con deviazione del becco superiore piů spesso a
destra.
Quest’anomalia puň essere scoperta giŕ a partire dal
9° giorno d’incubazione, ma la sua azione letale sull’embrione non si
rende manifesta se non a partire dal 18° giorno.
2.8. Esencefalo ereditario
|
Esencefalo
ereditario |
Ereditarietŕ
recessiva con bassa penetranza? Poligenico?
Gruppo di associazione sconosciuto
Sono stati descritti vari gradi di questa situazione,
caratterizzata da un’erniazione
del cervello per una schisi del cranio in sede frontoparietale,
dalla quale il cervello fuoriesce. A causa del cannibalismo,
questa situazione puň essere letale dopo pochi giorni dalla nascita. Se i
soggetti vengono allevati separatamente, possono raggiungere l’etŕ adulta a
prezzo di un particolare impegno da parte dell’allevatore. Vengono colpiti
maschi e femmine in percentuale identica ed č stata invocata un’ereditarietŕ
recessiva con penetranza molto bassa. Potrebbe tuttavia trattarsi di un’ereditarietŕ
su base poligenica.
2.9. Testa da passerotto
|
sph
- sparrow head |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Si tratta di una situazione letale tardiva, in cui i
pulcini sono normali alla nascita, ma progressivamente manifestano le
caratteristiche alterazioni a partire dalla nona settimana di vita. Si
instaura un progressivo accorciamento del becco e delle ossa del capo che
conferiscono alla testa un tipico aspetto da passerotto. I soggetti vanno
incontro a morte tra gli 8 e i 15 mesi d’etŕ, e nessuno riesce a
raggiungere la maturitŕ sessuale.
2.10. Naso camuso
|
sno
- snub nose |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Il naso camuso non č altro che il naso schiacciato, forse
derivato da muso con ca- peggiorativo, oppure dal latino camus,i,
che significa museruola. La caratteristica principale determinata da questa
mutazione č una testa tozza e larga, dotata di un becco accorciato, con vari
gradi di becco da parrocchetto. I casi estremi mostrano becco incrociato e
taglia ridotta per difficoltŕ ad alimentarsi.
2.11. Palatoschisi
|
Palatoschisi |
Dovuta
a 2-3 loci recessivi
Gruppo di associazione sconosciuto
Si tratta di un difetto geneticamente complesso, in cui il
palato puň essere diviso sia da un solo lato che da ambedue. Il ceppo di
Leghorn Minnesota 420 č stato la sorgente iniziale di quest’anomalia,
presente nel 17% dei soggetti. Il sospetto di un’origine genetica č sorto
sottoponendo le femmine a una dieta deficiente in riboflavina: esse erano in
grado di incrementare l’incidenza della palatoschisi fino al 47,4% nel giro
di due anni. Non esiste una correlazione coi cromosomi sessuali e l’espressione
č identica nei due sessi. I dati concorderebbero con il seguente modello
genetico: intervento
di 3 loci recessivi durante un’alimentazione normale della madre, mentre in
corso di carenza di riboflavina interverrebbero 2 loci recessivi.
La penetranza sarebbe dovuta a ulteriori 3-4 loci minori. Nell’incremento
percentuale osservato a carico del gruppo tenuto a dieta carente, si
verificherebbe un’eliminazione dei geni aggiunti che controllano la
penetranza.
2.12. Fori al palato
|
pt
- palatal pits |
Autosomico
recessivo
Gruppo di associazione sconosciuto
La descrizione di fori di varie dimensioni a carico del
palato risalgono sempre al ceppo Minnesota 420, ma pare che i due
difetti del palato siano geneticamente correlati.
3. Collo
3.1. Collo torto
|
crn
- crooked neck |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Fu
descritto in Livorno perniciate, i cui pulcini, inizialmente normali, a metŕ
dello sviluppo somatico presentavano un collo torto per lo piů girato a
destra, con incidenza uguale nei due sessi. Lo sviluppo somatico č ovviamente
ritardato per problemi dovuti all’alimentazione, le femmine depongono poche
uova e i galli non riescono ad accoppiarsi.
L’intimo meccanismo dell’anomalia pare consista in un
arresto dello sviluppo oppure in un’errata distribuzione di uno o piů
legamenti della colonna vertebrale.
4. Tronco
4.1. Sterno curvo
|
Sterno
curvo |
Ereditarietŕ
di tipo dominante con interazione ambientale
Gruppo di associazione sconosciuto
Negli
uccelli lo sterno č molto sviluppato, specie nei buoni volatori, in quanto
offre l’inserzione ai muscoli deputati al volo. La superficie esterna dello
sterno, o superficie ventrale, č irregolare, soprattutto per la presenza di
una salienza lungo la linea mediana, la carena.
Abitualmente
quest’osso č rettilineo, ma puň presentare delle curvature, per lo piů
verso destra, e vari gradi di concavitŕ. Sono stati invocati numerosi fattori
per giustificare questa deformazione: oltre a una suscettibilitŕ genetica,
potrebbero entrare in gioco il sesso, l’etŕ alla quale il soggetto inizia
ad appollaiarsi, la larghezza dei posatoi. Esistono infatti studiosi che
escludono fattori genetici e attribuiscono il difetto a soli fattori ambientali, altri che hanno potuto dimostrare un’influenza
genetica
attraverso l’incidenza di sterno torto a carico dell’80% della progenie di
genitori dotati della stessa anomalia.
Se
i sostenitori dell’origine genetica vogliono corroborare la loro convinzione,
posso aggiungere che nell’uomo č frequente il riscontro del cosiddetto pectus
excavatum,
causa di errate interpretazioni radiografiche della volumetria cardiaca, in quanto a prima
vista il cuore puň sembrare ingrandito a carico del settore sinistro. Se al pectus
excavatum, rientranza piů o meno spiccata della parte inferiore dell’osso
tanto da permettere la raccolta anche di una bella manciata d’acqua, si
associa la sindrome della schiena dritta, allora al cuore viene impresso un
movimento rotatorio, per cui la parte dx si sposta verso il lato opposto dando
una falsa immagine radiologica di cardiomegalia.
Orbene,
anche se non si puň mettere la mano sul fuoco, basta guardare il torace di
uno dei genitori e troveremo la stessa anomalia. Un’anomalia non dev’essere
giudicata in modo forzatamente negativo. Infatti, nella donna, la rientranza
dello sterno č in grado di conferire una particolare disposizione dei seni, i
quali, essendo slanciati in avanti, e risultando salienti rispetto al centro
del torace parzialmente infossato, suscitano una sensualitŕ del tutto
particolare. Lo stesso puň valere nel caso di certe anomalie del pollo, che,
se non sono orripilanti, potrebbero rappresentare delle particolaritŕ che
solo il gusto estetico del giudice puň mettere in giusta luce. Forse che un
Collo Nudo, in assoluto, č qualcosa di gratificante? Tutti sappiamo perň che
la bruttezza di certi divi č ineguagliabile e accattivante, irrepetibile.
Basti pensare ai dentoni di Gino Bramieri, che quando era rotondo poco mancava
somigliasse a un tricheco.
Un’altra
cosa č il petto
da calzolaio,
frequente in passato quando i giovani si avviavano precocemente a questa
professione. A forza di premere con le calzature sull’osso ancora malleabile,
veniva a determinarsi una rientranza del torace in corrispondenza dello
sterno. Logicamente non sarŕ successo a tutti i ragazzi di incorrere nella
deformazione. Tralasciando ovviamente l’entitŕ della pressione esercitata,
parametro difficilmente quantificabile, si puň supporre che la deformabilitŕ
ossea sia un qualcosa di individuale, dipendente ovviamente da fattori
genetici.
Torniamo
al pollo. I sostenitori dell’origine ambientale favorita da una suscettibilitŕ
genetica hanno
potuto dimostrare che a 6 mesi d’etŕ l’incidenza si abbassa dal 92% all’11%
se si sono approntati posatoi larghi 2,5 cm all’etŕ di 12 settimane invece
di ricorrere precocemente al loro impiego a 4 settimane di vita. Tra i maschi
geneticamente non suscettibili, esposti a condizioni estreme favorenti la
deformitŕ sternale, solo il 3% ha mostrato di venirne colpito.
Dati piů recenti riferiti da Knize (1983) paiono dimostrare che l’ereditarietŕ dello sterno deforme č di
tipo dominante,
e che i fattori ambientali interagiscono con quello genetico. Giŕ Warren nel
1937 aveva affermato che si tratta di una specifica debolezza dello sterno e
non di un suo disturbo d’ossificazione.
4.2. Senza posteriore dominante
|
Rp
- dominant rumplessness |
Autosomico
dominante
Gruppo di associazione sconosciuto
Il
numero di elementi che compongono la colonna vertebrale č variabile non solo
da un uccello all’altro, ma anche tra individui appartenenti alla stessa
specie. La distinzione delle vertebre in cervicali, toraciche, lombari,
sacrali e coccigee č pure soggetta a variazioni per mancanza di accordo fra
gli studiosi.
Per
le vertebre dell’ultimo tratto dello scheletro assile del pollo ci atterremo
ai dati contenuti in Avian anatomy integument di Lucas e Stettenheim (1972).
Il sinsacro del
pollo č costituito da un totale di 15 vertebre
tra loro fuse in un osso unico detto appunto sinsacro e che sono cosě
suddivise: 4
lombari, 5
sacrali, 6
caudali.
Il
sinsacro appare come una struttura allungata e leggermente concava
ventralmente e che ai lati si articola con le ossa iliache del bacino. Dopo la
sesta vertebra caudale del sinsacro la colonna si continua nelle vertebre
coccigee, che sono esterne al sinsacro e mobili.
Queste
vertebre coccigee o caudali si suddividono in 6 vertebre
caudali e un
pigostilo. Il
pigostilo, un triangolo appiattito trasversalmente e terminante con un apice,
č l’ultima vertebra e deriverebbe dalla fusione di 6 vertebre terminali.
Sul pigostilo poggia la ghiandola uropigetica o uropigio![]() .
.
Dunn
e Landauer (1925) e Landauer (1928) si dedicarono allo studio della morfologia e
dell’ereditarietŕ della mancanza di posteriore nel pollo e giunsero alla
conclusione che questa situazione anatomica puň essere suddivisa in:
completa, intermedia, accidentale.
Esteriormente
la forma completa e quella accidentale apparivano essere la stessa cosa: i
soggetti avevano la parte posteriore del corpo arrotondata ed erano privi sia
di uropigio che di piume caudali. Ma, a un esame delle vertebre, le due forme
presentavano delle differenze.
La
forma
completa era
caratterizzata dalla mancanza di tutte le vertebre del pigostilo eccetto le
ultime due, di tutte le vertebre caudali libere e di una o due vertebre della
parte centrale delle vertebre caudali del sinsacro.
La
forma
accidentale
differiva da quella completa per il fatto che tutto quanto il pigostilo era
assente e che erano le ultime due vertebre caudali del sinsacro a essere
mancanti, invece di una o due della parte centrale.
La
forma
intermedia
differiva dalle altre due per il fatto che tutte le vertebre caudali libere
erano presenti, anche se fuse insieme in modo irregolare; come nella forma
completa, il pigostilo era assente eccetto le sue due ultime vertebre e
mancavano uno o due elementi della parte centrale delle vertebre caudali del
sinsacro. Da notare che nel 25% dei soggetti con forma intermedia era presente un uropigio
rudimentale nonché un numero variabile da 2
a 17 di
piume caudali.
Davenport
(1906, 1909) fu
uno dei primi a riferire studi genetici sulla mancanza di posteriore nel
pollo, concludendo che era coinvolto un gene dominante. Dunn (1925)
verificň lo stato
dominante del tipo completo, giungendo pure alla conclusione che la forma
accidentale non era di origine genetica. Dunn e Landauer (1936) assegnarono
al gene della forma completa il simbolo Rp.
La
base genetica della forma intermedia - o modificata - della mancanza di
posteriore venne pure studiata da Dunn e Landauer (1934, 1936) i
quali giunsero alla conclusione che i soggetti affetti dalla forma intermedia
potevano essere sia omozigoti che eterozigoti per Rp, con
l’intervento di geni modificatori multipli e recessivi portati dai soggetti
normali, i quali grazie a questi geni modificatori tendono a sopprimere
l’anormale sviluppo del posteriore.
La
mancanza di posteriore puň giŕ essere notata all’atto della schiusa. Nella
vita adulta per semplici ragioni meccaniche puň comportare una diminuzione
della feconditŕ: infatti durante l’accoppiamento un gallo normale si
mantiene in equilibrio grazie alla coda che gli serve da bilanciere, mentre ciň
non č possibile nei galli scodati nei quali la copula risulta incompleta. Un
handicap ulteriore č rappresentato da un folto piumino che puň circondare la
cloaca e che vale la pena rimuovere in prossimitŕ del periodo riproduttivo.
Tuttavia l’assenza di coda pare vantaggiosa in quanto consentirebbe di
sfuggire alle insidie della volpe.
A
parte la feconditŕ minore per ragioni meccaniche, pare che i soggetti
portatori di questo gene siano dotati di una piů bassa sopravvivenza, specie nei primi due mesi di
vita. Anche le
percentuali di schiusa sono ridotte: la proporzione degli embrioni portatori
di Rp morti tra il 17° e il 21° giorno d’incubazione č spesso piů alta di quanto ci si
aspetterebbe, e minore č anche il numero di pulcini che riescono a nascere.
Questi rilievi sono validi per ambedue i fenotipi.
La
mancanza di posteriore puň venir indotta anche sperimentalmente: Landauer (1945)
č riuscito a
riprodurre questa situazione anatomica nel 42% degli embrioni sviluppati da uova nel cui tuorlo era stata
iniettata insulina prima dell’inizio dell’incubazione. Altre modalitŕ
capaci di indurre l’anomalia sono le brusche variazioni di temperatura
dell’incubatrice durante la prima settimana, oppure lo scuotimento meccanico
delle uova prima di incubarle; con ambedue queste tecniche l’incidenza
dell’anomalia č pari a circa il 7%.
In
passato questa particolaritŕ anatomica fece attribuire al pollo il nome di Gallus
ecaudatus e forse uno dei primi a descriverlo, piů che
succintamente, č stato Ulisse Aldrovandi, che ne parla nel libro XIV
del II
volume della sua Ornithologia
dedicandogli il VII capitolo: De Gallo et Gallina ex Persia.

|
Quem vero
Persicum Gallum appellant, et quem hic depinximus, a nostratibus in eo
potissimum differt, quod cauda careat, caetera simillimus existit.
Crista tamen veluti caudam obtinet. Erat autem totus niger lineis
luteis conspersus: Alarum remiges principio albae erant, caetera atrae:
pedes cinerei: Gallina quoad formam habitumve nostratis erat similis:
colore a mare diversissimo, unde tam in his, quam in illis coloris
diversitatem vilipendendam arbitror. Erat autem tota coloris
ferruginei, tribus pennis remigum exceptis, quae nigrae erant. Crista, si cristae maris compares longe erat quam in
illo minor. |
Quel gallo che chiamano
persiano e che qui ho raffigurato differisce soprattutto dai nostri per il fatto di essere senza coda,
mentre per il resto č molto simile. Tuttavia la cresta possiede come una
coda. Era tutto nero e spruzzato di linee gialle; le remiganti erano
bianche all’apice e per il resto nere; le zampe erano color cenere.
La gallina era simile a quelle nostrane per forma o corporatura; di
colore era molto diversa dal maschio, per cui ritengo che sia nei
polli nostrani che in questi polli la differenza di colore č di
scarsa importanza. Era tutta quanta color ruggine, eccetto tre
remiganti che erano nere. La cresta, se paragonata a quella del
maschio, era molto piů piccola. |
Sempre
in passato, secondo Dunn (1925) l’assenza di posteriore č stata una caratteristica
presente in Combattenti nani e nella Polish scodata. Ora č un tratto
dell’Araucana, dell’Ardennese senza coda, della Kaul e di certe Barbute
Belghe come la Grubbe.
4.3. Senza posteriore recessivo
|
rp-2
- recessive rumplessness |
Autosomico
recessivo con bassa penetranza
Gruppo di associazione sconosciuto
Questo
gene ha
una bassa penetranza e un ampio grado d’espressivitŕ, influenzata da geni
modificatori, alcuni dei quali legati al sesso. Fu descritto in un ceppo di
Leghorn bianca.
Presso
gli allevatori quest’anomalia č nota come schiena da scarafaggio.
Le caratteristiche anatomiche sono simili a quelle della forma dominante:
pigostilo rudimentale, vertebre coccigee fuse in una protuberanza ossea. In
alcuni casi ambedue queste strutture ossee sono assenti. A seconda del grado
di interessamento scheletrico, l’uropigio![]() e le timoniere sono assenti oppure
č presente un abbozzo di ghiandola con timoniere cascanti e in numero ridotto.
e le timoniere sono assenti oppure
č presente un abbozzo di ghiandola con timoniere cascanti e in numero ridotto.
Quest’anomalia si distingue anatomicamente dalla forma
dominante per il fatto che le vertebre delle coda sono compresse lateralmente e dotate di una
curvatura rivolta verso il basso, e che il sinsacro č alterato, essendo talora
cifoscoliotico. I maschi presentano anomalie di grado piů marcato rispetto
alle femmine. L’alterazione č giŕ diagnosticabile al 4°-5° giorno d’incubazione,
ed esiste una correlazione tra grado di riduzione della coda e precocitŕ d’insorgenza.
Schiudibilitŕ e vitalitŕ sono ridotte come nella forma dominante.
4.4. Coste soprannumerarie
|
Coste
soprannumerarie |
Poligenico,
con un gene maggiore e numerosi modificatori, alcuni legati al sesso
Gruppo di associazione sconosciuto
Abitualmente
il pollo possiede 7 paia di coste: le prime due coppie sono dette fluttuanti in
quanto sono incomplete e non si congiungono allo sterno, le successive 4
paia si articolano con lo sterno, l’ultimo paio, composto dalla 7Ş costa, non si inserisce direttamente allo sterno bensě sulla sesta
costa, per cui la 7Ş
costa č detta costa falsa. Č stato descritto un 8°
paio di coste nella
Leghorn e queste coste soprannumerarie avevano le caratteristiche di coste
false, come quelle del settimo paio. L’incidenza dell’anomalia sembra
essere maggiore nel maschio. Č stata proposta un’ereditŕ poligenica in cui č presente un gene maggiore e numerosi geni
modificatori, alcuni dei quali legati al sesso.
5. Ali
5.1. Assenza di ali legata al sesso
|
wl - wingless |
Legato
al sesso, recessivo
Gruppo di associazione V - cromosoma Z
Questo
gene č legato ai loci S e K. I pulcini senz’ali
sono davvero unici, e i primi descritti erano tutte femmine in apparenza
normali, salvo l’assenza di ali. Nella discendenza di successivi incroci
comparvero l’assenza di ali e vari gradi di assenza di zampe, sino alla
mancanza dei 4 arti, detta amelěa. A questi casi estremi si
contrappongono quelli con penetranza nulla, cioč minore dell’1%,
rappresentati da soggetti con ali cadenti, remiganti anormali e talora falangi
soprannumerarie. Infatti l’espressione di questa mutazione č molto
variabile, ma in tutti i casi si ha l’interessamento delle ali, con organi
interni normali, scarso sviluppo della muscolatura del petto, occhi
lacrimevoli nei soggetti privi di ali.
5.2. Assenza di ali autosomica
|
wg-3
- wingless |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Citiamo
per completezza anche questa mutazione, riferita da Landauer nel 1973, senza
ulteriori dettagli a disposizione. Anatomicamente č del tutto simile alla
stessa anomalia legata al sesso.
6. Gambe
6.1. Gambe da bassotto, carponi
|
Cp
- creeper |
Autosomico
incompletamente dominante
Gruppo di associazione I - cromosoma 2
Appartiene
al 1° gruppo di linkage e dista solo 0,4 unitŕ crossover dal gene della
cresta a rosa R. Pertanto, una razza dotata del gene Cp mostra
una stretta associazione tra la mutazione zampe corte e cresta semplice
- che č il contrario della cresta a rosa - e che č dovuta al selvatico r+.
In inglese
to creep significa strisciare. Creeper equivale a Krüper,
termine che in basso tedesco corrisponde al tedesco Kriecher,
strisciante. Fra tutte le mutazioni che
interessano le appendici scheletriche, questo gene ha ricevuto l’attenzione
maggiore da parte degli studiosi, ed č presente in varie parti del mondo, in
quanto č una caratteristica distintiva di alcune razze, come Jitokko e
Nagasaki in Giappone, Courtes Pattes in Francia, Scot Dumpy - Scozzese
tarchiata - nel Regno Unito, Krüper in Germania. La Courtes-Pattes,
completamente scomparsa da alcuni anni, fu ricreata da Périquet a partire
dalla tedesca Krüper.
Giŕ
dal 1925 Cutler si era accorto che gli adulti creeper erano sempre
eterozigoti per un gene che in seguito si dimostrň essere autosomico nonché
letale se omozigote, per cui gli embrioni morivano intorno al 4° giorno. La
morte precoce pare sia dovuta a un’inibizione dello sviluppo del sistema
vascolare.
Pertanto non
esistono dei creepers puri omozigoti,
e quando due soggetti in regola con lo standard vengono accoppiati, Ľ delle
loro uova non schiude, il 50% riesce a schiudere e darŕ luogo a soggetti
eterozigoti per Cp, e Ľ dei pulcini sarŕ omozigote per cp+, pertanto con arti inferiori normali.
Questi
ultimi soggetti non sono da scartare, perché, a parte i tarsi di lunghezza
normale, portano con sé tutte le altre caratteristiche di razza e possono
essere accoppiati con soggetti a zampe corte. Cosě facendo tutte le uova
riescono a schiudere e la percentuale di soggetti a zampe corte
corrisponderŕ al 50%. Pochi sono gli omozigoti che sfuggono alla letalitŕ
precoce e che riescono a nascere. Sono tuttavia destinati a soccombere poco
dopo la nascita.
Posso
dire che mi capita spesso di osservare dei soggetti di Nagasaki arricciata i quali
rimangono piccolini piccolini. Ovviamente in un allevamento amatoriale č
difficile trarre dei dati scientificamente probanti, tuttavia credo non si
possa escludere che talora certi omozigoti riescano a sopravvivere per un
periodo abbastanza lungo. Ricordo che uno di questi scriccioli era vivacissimo
e non cresceva, sembrava sempre appena uscito dall’uovo pur vivendo la vita
dei genitori e dei fratelli. Venne un forte acquazzone e riuscii a salvarlo
dalla morte per raffreddamento. Una seconda volta fallii e lo persi per sempre.

Fig. X. 5 - Spennacchiona e Spennacchione
Posseggo
una coppia di Nagasaki arricciata pied, lui con la cresta pendula,
veramente esagerata rispetto al soma, lei buona fetatrice
[2]
e buona
covatrice. Lui, lo spennacchione, fu salvato quasi abbrustolendolo sulla stufa
durante una grave crisi dispnoica da tracheite, dopo di che ricevette un
cappotto ricavato da una calza di lana, continuň a vivere in casa, si
impiumň, dopo due mesi smise il soprabito, ora vive felicemente con la sua
dolce metŕ. Se saranno capaci di superare anche quest’inverno, metterň a
cova le loro uova e, salvo una sterilitŕ misconosciuta, vorrň vedere cosa ne
esce. Ambedue sono minuti, e so benissimo che il mancato incremento corporeo
puň dipendere da molteplici fattori, anche se nei miei Chabos pare un evento
troppo frequente. Conforta questa mia ipotesi l’affermazione di Fujio &
Shibuya, che hanno trovato una differente espressivitŕ dell’azione letale
di Cp, la quale dipende dallo sfondo genetico sul quale si esplica,
imputando la riduzione della mortalitŕ a geni autosomici dominanti, senza
poter escludere un meccanismo abbastanza strano, secondo cui parrebbe che
nelle prime fasi del periodo depositivo la madre inglobi nel citoplasma dell’uovo
una qualche sostanza capace di abbassare l’effetto precocemente letale di Cp.
Ma Landauer, dal canto suo, ottenne lo stesso risultato, triplicando
addirittura gli embrioni: incubň le uova piů tardive per le prime 24 ore a
35,5°C anziché a 37,2-37,7°C abituali. Lo stato Cp/Cp era ugualmente
attivo, e anche se ritardava lo sviluppo, agli embrioni era concesso di
procedere ulteriormente nella crescita.

Fig. X. 6 - Spennacchiona che cova

Fig. X. 7 - Spennacchiona madre felice
Quest’anomalia
appartiene alle malformazioni scheletriche che vanno sotto il nome di acondroplasia
o condrodistrofia.
Tutte le ossa lunghe degli arti inferiori sono accorciate, piů nei maschi che
nelle femmine. Stando a Landauer l’osso piů colpito č il tarsometatarso.
Secondo i dati di Stevens si tratta di un prevalente interessamento della
tibia, affiancata da una fibula lunga uguale e piů spessa della norma. L’embrione
eterozigote puň essere riconosciuto giŕ dal 6° giorno d’incubazione.
Studi
dettagliati sull’osteogenesi embrionale, nei pulcini in crescita e negli
adulti, hanno mostrato che si tratta di un disturbo dell’ossificazione encondrale,
quella che in condizioni normali permette all’osso di allungarsi
progressivamente per intervento della cartilagine. Mentre nel creeper
questo processo č disturbato, il processo di ossificazione che origina dal
periostio si svolge in modo normale, per cui le ossa lunghe diventano spesse e
tozze.
La
letalitŕ da Cp non si limita ai soli omozigoti, in quanto il gene si
dimostra letale anche per una quota degli eterozigoti valutata intorno al 5%.
A questo si aggiunga una letalitŕ embrionale ancora del 5% per quelle uova
deposte da femmine creeper che presentano una variabilitŕ a carico
dello spessore del guscio.
Esiste
un’altra iettatura per i creepers che hanno avuto la fortuna di
essere eterozigoti: essi sono piů suscettibili a una carenza di vitamina D e
agli effetti tossici del selenio. Come abbiamo giŕ detto, non facciamo l’idiozia di sbarazzarci dei
soggetti a gambe normali, specialmente se dotati di caratteristiche che vale
la pena mantenere. Essi servono egregiamente per abbattere la letalitŕ
embrionale, in quanto, incrociati con soggetti da esposizione, daranno una
progenie tutta vitale, anche se solo la metŕ sarŕ secondo lo standard.
Oltretutto, nasceranno molti piů bassotti da questo incrocio che accoppiando
due eterozigoti. Lo stesso discorso č applicabile al caso della Cornish,
anche se il gene in causa č differente.
Il
gene Cp č responsabile dell’accorciamento e dell’ispessimento dei
tarsi in modo variabile, per cui, per giustificare la variabilitŕ del
fenotipo, bisogna invocare o una capacitŕ estrema di raccorciare dovuta al
gene Cp, oppure dei geni modificatori. Non si tratta di un’azione
dose dipendente come nel caso del gene B del barrato legato al sesso, in quanto gli omozigoti per Cp
vanno incontro a morte quasi certa intorno al 4° giorno d’incubazione.
Si postula l’intervento di geni
modificatori in
base alla constatazione che, per variare la lunghezza dei tarsi, per
proporzionare lunghezza delle ossa e delle cosce, per conferire alle ossa il
giusto grado di rotonditŕ, per non determinare stonature tra spessore dei
tarsi e lunghezza delle dita, debbono essere per forza in gioco dei geni di
cui conosciamo poco o nulla. Pertanto l’allevatore non deve far altro che
selezionare e accoppiare i soggetti con le caratteristiche volute,
incanalando cosě nel corredo tutti i geni necessari al raggiungimento dello
scopo prefisso.
Chiudiamo questo paragrafo con una digressione genetica dell'uomo e del cane che possono presentare la stessa patologia osservabile nel pollo. Il nano di un circo equestre č tale in quanto affetto da nanismo acondroplasico dovuto a un gene dominante posto sul cromosoma 4 (fibroblast growth factor receptor-3 gene, acronimo FGFR3) letale per il feto o subito dopo la nascita se omozigote, quindi letale come nel pollo. Nel noto cane bassotto tedesco, detto Dachshund o cane tasso, il gene FGFR3 si trova sul cromosoma 3, dominante nel Basset e recessivo nel Cocker Spaniel. Grazie al Dr Fabrizio Bevilacqua, veterinario a Moncalvo (AT), possiamo aggiungere che, per quanto finora č dato sapere, non si puň supporre nei cani bassotti una mortalitŕ embrionale precoce su base genetica in analogia a quanto dimostrato nel pollo. Ecco la sua e-mail dell'8 febbraio 2008.
Ciao Elio,
Come accennato al telefono, ti confermo che, nella mia casistica personale e
nella bibliografia di mia conoscenza, nel Bassotto la discrepanza tra il
conteggio delle camere gestazionali effettuato ecograficamente (a 23/28 giorni
post-accoppiamento) e il numero di cuccioli partoriti non si discosta rispetto
ad altre razze; inoltre, il numero medio di cuccioli nati in questa razza č
in linea a quanto ci si aspetterebbe per soggetti di tale mole. Purtroppo, ad
oggi, non disponiamo di tecnologie che ci permettano di valutare piů
precocemente lo stato gravidico e il numero di embrioni e pertanto non
possiamo supporre una mortalitŕ embrionale precoce su base genetica in
analogia a quanto dimostrato nel pollo.
Con la speranza di esserti stato utile, ti saluto cordialmente.
Dr
Fabrizio Bevilacqua - DVM - Moncalvo (AT)
8 febbraio 2008
Gallina
bassotta bolognese
del senatore Fulvio Antonio Marescalchi
Riassunto
Il
testo completo della ricerca di genetica storica
si trova nel lessico alla voce Gallina bassotta bolognese![]()

La gallina monstrosa del senatore bolognese Fulvio Antonio
Marescalchi raffigurata a pagina 562 del Monstrorum historia
di Ulisse Aldrovandi, pubblicato postumo da Bartolomeo Ambrosini![]() , nel 1642. Questa gallina – verosimilmente una creeper
– forse č il primo caso di tale mutazione genetica a fare la sua comparsa
in letteratura. Essa fu inserita nel capitolo dedicato ai soggetti dotati di
piedi mostruosi - Multiplicatio pedum
in foetibus avium - anche se dotata di piedi normali. Ma per l'anomala
struttura del tarsometatarso il suo incedere era del tutto anormale.
, nel 1642. Questa gallina – verosimilmente una creeper
– forse č il primo caso di tale mutazione genetica a fare la sua comparsa
in letteratura. Essa fu inserita nel capitolo dedicato ai soggetti dotati di
piedi mostruosi - Multiplicatio pedum
in foetibus avium - anche se dotata di piedi normali. Ma per l'anomala
struttura del tarsometatarso il suo incedere era del tutto anormale.
Se analizziamo la letteratura antica a partire da Aristotele, Columella e Plinio, per finire con quella piů recente di Longolius, Gessner e Aldrovandi, non riusciamo a identificare un pollo con le caratteristiche anatomiche causate da uno dei geni dell'acondroplasia (Cp o Mp), capaci di determinare uno spiccato accorciamento delle zampe. Da parte di questi autori vengono citati dei polli nani, ma sono tali non per l'acondroplasia, bensě a causa di una riduzione della mole corporea su base genetica. Un trabocchetto linguistico potrebbe essere rappresentato dal termine Däsehünle riferito da Gessner, traducibile con gallinetta tasso, un trabocchetto in quanto il cane bassotto viene detto cane tasso, Dachshund, dotato di un corredo genetico che determina l'acondroplasia. Ma il trabocchetto teso dal vocabolo Däsehünle viene dissipato dalle parole dello stesso Gessner che inequivocabilmente identifica la Däsehünle con una gallina di corporatura piccola: "Un nostro amico spagnolo chiama in spagnolo la gallina Adriatica gallina enana. Certamente in quanto č di corporatura nana e piccola, e tale razza presso di noi in Svizzera sento dire che viene chiamata Schotthennen, in altri posti Erdhennle, altrove Däsehünle." La prima descrizione di una gallina affetta verosimilmente da acondroplasia č indirettamente dovuta ad Aldrovandi, in modo diretto al suo successore Bartolomeo Ambrosini che inserě questa gallina nel Monstrorum historia di Aldrovandi, edito postumo nel 1642. Sotto il profilo anatomico la descrizione di Ambrosini č dettagliata e presenta un aggancio quasi indiscutibile con la letalitŕ embrionale dovuta all'omozigosi per il gene dell'acondroplasia, nonché un aggancio puramente iconografico con la cresta semplice il cui gene risiede sul cromosoma 2 tanto come Cp e Mp. Non sappiamo quando una di queste due mutazioni genetiche raggiunse l'Europa. Č assai verosimile trattarsi del XVI secolo. I Tedeschi non chiamarono gallinetta tasso questo pollo, bensě Krüper, strisciante, e questa sarebbe una riprova che Däsehünle di Gessner non equivale a Krüper. Attraverso una citazione di Buffon da lui mistificata possiamo presumere che la mutazione ebbe origine in Cambogia, quindi passň nelle Filippine da dove nel XVI secolo gli Spagnoli la portarono in Europa attraverso il Pacifico, il Messico e poi l'Atlantico. Non esistono menzioni di galline paticortas in Spagna nel XVI secolo. Padre Bernabé Cobo - o chi per esso - scrisse che gli Spagnoli portarono in Messico dalle Filippine una gallina, ma non venne descritta nelle sue caratteristiche fenotipiche. Si trattava verosimilmente della paticortas, della dumpy, che grazie agli Spagnoli oltre al nord Europa e alla Scozia raggiunse anche Bologna.
6.2. Membra corte, ametapodia
|
Mp
- ametapodia |
Autosomico
incompletamente dominante
Gruppo di associazione I - cromosoma 2
I
soggetti affetti da quest’anomalia sono eterozigoti, in quanto la situazione
omozigote č letale per l’embrione, con un picco di mortalitŕ al 17°
giorno. A differenza degli eterozigoti per Cp, non č presente un
incremento della mortalitŕ embrionale e una riduzione della schiusa. Anche
per questo gene esiste un rapporto d’associazione col gene della cresta a
rosa, che perň č di 16 unitŕ crossover, e Mp giace sicuramente in un locus
distinto da quello occupato da Cp.
Sotto molteplici aspetti questa mutazione somiglia alla creeper,
anche se i soggetti hanno gambe molto piů accorciate. Interessa
principalmente gli arti inferiori e in modo meno spiccato anche le ali; il
soma non presenta alterazioni ulteriori. Il tarsometatarso e il carpometacarpo
sono assenti, ma tutte le dita sono pressoché normali. Con l’invecchiamento,
varie ossa della zampa e delle dita tendono a fondersi, col risultato di una
perdita dell’articolazione. I soggetti affetti da ametapodia possono
riprodursi, ma il loro allevamento richiede una cura estrema in quanto hanno
problemi deambulatori: camminano zoppicando e non sono in grado di mantenere
la stazione eretta, in quanto non possono servirsi delle dita come appoggio.
6.3. Letale della Cornish
|
Cl - Cornish
lethal |
Autosomico
incompletamente dominante
Gruppo di associazione sconosciuto
Se
omozigote, si dimostra letale al termine dell’incubazione, se eterozigote
contribuisce ad accorciare le estremitŕ. La Cornish č caratterizzata da un
corpo molto allargato che poggia su zampe corte e robuste; anche la testa č
breve e larga. Questi tracagnotti sono dotati di tibiotarso e di
tarsometatarso spiccatamente corti, con fibula completamente sviluppata cosě
da estendersi fino alla parte terminale e inferiore della tibia, dove si fonde
con l’epifisi di quest’osso. Spesso sono accorciate anche le ossa delle
dita.
L’effetto
di questo gene deriva dall’influsso di geni multipli, dei quali uno solo o
parecchi si comportano come dominanti; uno di questi geni modificatori č
letale se omozigote, e non tutti i ceppi di Cornish ne sono dotati; pare sia
presente solo in quei casi di arti inferiori estremamente accorciati. Tra
tutti i geni che possono essere cointeressati in questo fenotipo, il simbolo Cl
č stato attribuito solo a quel gene dotato di azione letale.
Da notare che non sempre ciň che comporta un premio alle
esposizioni appaga le aspettative dell’allevatore, in quanto quei galli
Cornish estremamente tozzi e larghi danno spesso problemi di fertilitŕ.
Pertanto, ancora una volta bisogna far leva sia sul gusto estetico che sulle
capacitŕ dell’allevatore per ottenere la giusta miscela di geni che non
comprometta né la schiudibilitŕ né la capacitŕ di accoppiamento dei galli.
L’ideale sarebbe poter disporre di maschi non da primo premio ma dalle gambe
sufficientemente lunghe, nonché di femmine discretamente accovacciate.
6.4. Perosi su base genetica
|
pe
- perosis |
Autosomico
recessivo
Gruppo di associazione III - cromosoma 1
Perosi
deriva dal greco përós, sciancato, storpio. Come si č detto nel
paragrafo relativo alle influenze ambientali![]() sugli esseri viventi, gli animali differiscono geneticamente anche nei
confronti dei loro fabbisogni nutritivi. I polli Livorno possono accrescersi
con una razione che contiene solamente 30 parti per milione di manganese,
mentre Rhode Island e altre razze pesanti richiedono almeno 50 parti p.m. se
si vuole evitare la perosi,
cioč una condizione di osteocondrodistrofia in cui l’articolazione del
calcagno (quella tra tibia e tarsometatarso) si ingrossa provocando zoppia.
Questa perosi č su base alimentare cui sottende un substrato genetico
difficile da individuare, tant'č che abitualmente per perosi si intende una
malattia che colpisce i pulcini e i tacchinotti, caratterizzata dalla torsione
laterale della tibia o tibiotarso con rigonfiamento e arrossamento del
calcagno. Sembra sia originata da carenza di particolari sostanze (manganese,
colina, biotina, ecc.) e dalla somministrazione di una dieta particolarmente
ricca di mais e di sostanze proteiche.
sugli esseri viventi, gli animali differiscono geneticamente anche nei
confronti dei loro fabbisogni nutritivi. I polli Livorno possono accrescersi
con una razione che contiene solamente 30 parti per milione di manganese,
mentre Rhode Island e altre razze pesanti richiedono almeno 50 parti p.m. se
si vuole evitare la perosi,
cioč una condizione di osteocondrodistrofia in cui l’articolazione del
calcagno (quella tra tibia e tarsometatarso) si ingrossa provocando zoppia.
Questa perosi č su base alimentare cui sottende un substrato genetico
difficile da individuare, tant'č che abitualmente per perosi si intende una
malattia che colpisce i pulcini e i tacchinotti, caratterizzata dalla torsione
laterale della tibia o tibiotarso con rigonfiamento e arrossamento del
calcagno. Sembra sia originata da carenza di particolari sostanze (manganese,
colina, biotina, ecc.) e dalla somministrazione di una dieta particolarmente
ricca di mais e di sostanze proteiche.
Nel
1969 Somes ha studiato un tipo di perosi in una popolazione presa a caso e
mantenuta a dieta corretta per tutta la durata delle osservazioni. Si č
potuta dimostrare un’ereditarietŕ basata su un singolo gene recessivo con
una penetranza, negli omozigoti, pari al 74%.
I soggetti con perosi evidenziarono i sintomi fra le 3 e
le 5 settimane d’etŕ. L’espressione dell’anomalia era ampiamente
variabile: abitualmente la perosi iniziava con un ingrossamento dell’articolazione
del calcagno, seguito da vari gradi di torsione del tibiotarso e di
deviazione del tarsometatarso. Nei casi estremi l’arto rimaneva fissato in
estensione. L’interessamento era mono o bilaterale, e molto spesso uno o
ambedue gli arti impiegavano otto settimane per tornare alla normalitŕ.
6.5. Anomalia dell’articolazione tibio-tarsometatarsica
|
Anomalia
dell’articolazione tibio-tarsometatarsica |
Poligenico
Gruppo di associazione sconosciuto
Pare verosimile che alcune anomalie a carico degli arti
inferiori abbiano una base
poligenica. I
casi studiati da Serfontein & Payne nel 1934 avevano fatto la loro
comparsa fra le 3 e le 8 settimane d’etŕ. I primi sintomi sono lo squatting,
cioč l’accoccolarsi e lo star seduti alla turca, accompagnato da turgore e
dilatazione delle vene safene. Col progredire dell’affezione, la tibia e il
tarsometatarso vanno incontro a deviazione e ad incurvamento, cui si associa
spesso la fuoruscita del tendine del gastrocnemio dalla sua sede normale.
Talora i muscoli delle gambe perdono la capacitŕ di flettersi e rimangono
fissati in estensione. Pare che le razze pesanti siano piů spesso colpite da
quest’anomalia.
6.6. Zoppia
|
Zoppia |
Poligenico
Gruppo di associazione sconosciuto
Nel 1955 Morejohn ha riferito di una zoppia su base
ereditaria, in cui č implicato senz’altro piů di un singolo gene, che
comprometteva il bacino, le articolazioni delle gambe, le palpebre, queste
ultime corrugate nei soggetti piů colpiti. Questa mutazione č stata
rinvenuta in un ceppo di Cornish scura. Ambedue le articolazioni
tibio-tarsometatarsiche avevano una limitazione del movimento di flessione,
con graduale torsione e rotazione verso l’esterno, cui si associavano dita
arrotolate in flessione oppure in estensione. Le anomalie a carico del bacino
probabilmente erano secondarie all’azione muscolare rivolta a mantenere una
stazione corretta, accompagnata pur tuttavia da dita arricciate.
6.7. Perosi congenita
|
cop -
congenital perosis |
Autosomico
recessivo
Gruppo di associazione sconosciuto
I pulcini hanno le gambe rigirate e muoiono pochi giorni
dopo la schiusa. Questa mutazione sembrerebbe del tutto simile a quella
descritta da Somes nel 1970 e da Morgan nel 1963.
6.8. Discondroplasia tibiale
|
Discondroplasia
tibiale |
Poligenico
con modificatori legati alla dieta
Gruppo di associazione sconosciuto
Sheridan
(1978)
ha ipotizzato
trattarsi di un’anomalia dovuta in prevalenza a un gene recessivo legato al
sesso, col quale interferiscono sia componenti di origine materna sia geni
dominanti, da soli o in cooperazione, per cui siamo in presenza di un’anomalia
su base multifattoriale
alla quale non sono estranei fattori modificatori legati alla dieta.
Negli Stati Uniti quest’anomalia delle gambe č nota
come osteodistrofia focale, e si tratta di un’ipertrofia della cartilagine
della tibia e del tarsometatarso presente nei broilers, riscontata in diversi
Paesi. L’ipertrofia della cartilagine č di forma conica e si presenta a
carico del piatto tibiale, per cui si determina un ingrossamento del ginocchio
con zoppia. Sono state prodotte delle linee con bassa incidenza dovuta ai soli
fattori alimentari.
6.9. Zoppia congenita
|
coc - congenital
crippling |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Sono implicati anche geni modificatori. Questa zoppia č
stata descritta da Somes nel 1970 in soggetti nati da una serie di incroci tra
Plymouth Rock bianca con New Hampshire. L’anomalia non era isolata, ma gli
altri tratti segregavano in modo indipendente. Alla schiusa, una o ambedue le
gambe sono proiettate lateralmente, rendendo i movimenti del pulcino
estremamente difficoltosi, per cui la letalitŕ interviene per impossibilitŕ
ad alimentarsi.
6.10. Anomalia congenita delle gambe
|
cla
- congenital leg anomaly |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Č
in combutta con geni modificatori. Si tratta di un’anomalia riscontrata in
un ceppo di Plymouth Rock, giŕ evidente alla schiusa, capace di causare una
zoppia permanente in quanto la testa femorale non si articola con l’acetabolo.
Una o ambedue le gambe sono interessate e si presentano estese lateralmente,
anteriormente o superiormente. I soggetti colpiti erano in prevalenza maschi,
83%, e non sopravvissero fino all’etŕ matura.
6.11. Micromelia Hays
|
mm-H
- micromelia Hays |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Č una mutazione letale per una parte degli eterozigoti.
Il nome č legato alla descrizione fatta da Hays (1944) di
una mutazione letale nella RIR con segregazione di 54 pulcini anormali su un
totale di 172. Il fenotipo era uniforme: testa larga e appiattita, mascella e
mandibola ridotte notevolmente di volume con forma regolare, ossa lunghe di
gambe e ali molto accorciare e ispessite, con ossa delle gambe che tuttavia
erano dritte. I pulcini nati si dibattevano facendo perno sulla pancia a causa
della brevitŕ delle gambe. Di questi pulcini il 30% riuscě a venire alla
luce, morendo poi nel giro di una settimana.
6.12. Senza stinchi
|
shl
- shankless |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Questa
mutazione, che nessuno di noi avrŕ la ventura di vedere, viene riportata in
quanto č dovuta a uno dei meccanismi cromosomici capaci di causare una
mutazione: l’inversione
pericentrica,
che in questo caso č a carico del cromosoma 2, ottenuta coi raggi X.
Generalmente si esprime come autosomica recessiva. Questa regola fa un’eccezione
negli eterozigoti, che presentano un grado intermedio di accorciamento del
metacarpo.
Č stata descritta nel 1985 da Langhorst & Fechheimer
come anomalia che interessa unicamente gli arti: metacarpi malformati e di
dimensioni ridotte, assenza del tarsometatarso, tibia piů lunga, ossa
soprannumerarie a carico di 2°-3°-4° dito con fusione delle falangi
prossimali. Questa situazione genetica č per lo piů letale verso la fine
dell’incubazione o poco dopo la nascita, anche se talora alcuni soggetti
possono sopravvivere fino a riprodursi, con fertilitŕ del maschio non diversa
dalla norma.
Mutazioni a psicogenesi
umana
La manucodiata
Nei secoli passati non fu mai descritto nel pollo un fenotipo come quello della manucodiata, l’uccello del paradiso, Paradisea apoda, dove apoda significa senza zampe. Forse perché il pollo, vivendo al fianco di noi comuni mortali, ed essendo per definizione un ruspante, non si prestava a fantasticherie eccessive, come l’attribuirgli una mancanza di zampe.
Invece
una sfilza di uccelli delle Indie Orientali incappň in questa singolare
mutazione, superiore in assoluto a tutte quelle appena descritte nel pollo,
sfilza d’uccelli il cui prototipo fu appunto la Paradisea apoda,
cosě battezzata da Linneo nel 1760,
quasi a voler perpetuare nei secoli le leggende che l’uomo grazie alla sua
ineguagliabile fantasia aveva ricamato in proposito, sulla spinta e sulla
scia del molto lucroso basilisco![]() .
.
Tuttavia, come si puň dedurre dal naturalista inglese Alfred Russel Wallace (1823-1913), la classificazione di Linneo non voleva assolutamente perpetuare la leggenda nei secoli a venire, in quanto nel 1760 nessun esemplare integro di Paradisea apoda aveva mai messo piede in Europa, né alcunché di preciso era ufficialmente noto sul suo conto: “Down to 1760, when Linnaeus named the largest species, Paradisea apoda (the footless Paradise Bird), no perfect specimen had been seen in Europe, and absolutely nothing was known about them.” Ma, come vedremo tra poco, Wallace fa un'affermazione che giŕ nel 1669 veniva smentita da Georg Horst: infatti una, almeno una manucodiata integra, cioč fornita di zampe, aveva messo piede in Europa un bel po' prima del 1760. E perdipiů, non distante da dove visse e operň Linneo.

Paradisi avis vel Paradisea – Italice Manucodiata
da Icones avium omnium di Conrad Gessner (1560)
secondo il quale la manucodiata
proveniva nel Nuovo Mondo
forse perché nel XVI secolo le Molucche appartenevano al Nuovo Mondo.
In Historia Animalium III (1555) a pagina 612![]() specifica che dai
Maomettani era invece detta mamuco diata
specifica che dai
Maomettani era invece detta mamuco diata
e dŕ prova di
essere ben conscio che la Paradisea proveniva dalle Molucche.
Manucodiata
- anche manucco diatta e mamuco diatta - č la corruzione del
malese manuk dewata, l’uccello
di Dio. Da oltre 500
anni le penne di questo uccello e dei sui congeneri affascinano per la loro
bellezza Europei e non Europei, e giŕ molti secoli or sono le prime spoglie
di questi volatili venivano portate dalla Nuova Guinea in Cina e sulle isole
della Sonda. Le spoglie della manucodiata pare siano giunte in Europa con
l'unica superstite delle cinque navi che componevano la flotta di Magellano,
la Victoria, comandata da Elcano, e che grazie a lui riapprodň a Sanlúcar
de Barrameda il 6 settembre 1522,
in quanto Magellano si era spinto nelle Filippine e qui era stato ucciso nel
1521 sull’isola
di Mactan dagli indigeni, esasperati dagli esosi tributi. Tra i 18
superstiti della spedizione c’era Antonio Pigafetta (Vicenza ca. 1485 - ca. 1534)![]() , un gentiluomo vicentino che in qualitŕ
di criado, cioč di addetto alla
persona, seguiva il capo della spedizione, ed č a lui che si deve l’unica
testimonianza diretta di tutto il viaggio intrapreso da Magellano. Gli
indigeni delle isole Aru - arcipelago del Mar degli Arafura, tra le isole
Kai a ovest e la Nuova Guinea a est, amministrativamente compreso nella
provincia indonesiana delle Molucche - effettuavano sulla Paradisea apoda
tutta una serie di manipolazioni: dopo aver tolto le zampe, le ossa e le
interiora, lasciavano essiccare la pelle dell'animale, che rimaneva gonfia e
intatta, anche se vuota, con l'effetto di un'incredibile leggerezza.
Successivamente inserivano su questa pelle numerose variopinte penne
esotiche.
, un gentiluomo vicentino che in qualitŕ
di criado, cioč di addetto alla
persona, seguiva il capo della spedizione, ed č a lui che si deve l’unica
testimonianza diretta di tutto il viaggio intrapreso da Magellano. Gli
indigeni delle isole Aru - arcipelago del Mar degli Arafura, tra le isole
Kai a ovest e la Nuova Guinea a est, amministrativamente compreso nella
provincia indonesiana delle Molucche - effettuavano sulla Paradisea apoda
tutta una serie di manipolazioni: dopo aver tolto le zampe, le ossa e le
interiora, lasciavano essiccare la pelle dell'animale, che rimaneva gonfia e
intatta, anche se vuota, con l'effetto di un'incredibile leggerezza.
Successivamente inserivano su questa pelle numerose variopinte penne
esotiche.
Tra le meraviglie che i marinai della Victoria avevano a bordo c’erano alcuni esemplari di manucodiata. Venne subito rilevato il fatto straordinario che i corpi ben conservati non possedevano né piedi né ossa. Antonio Pigafetta, pochi anni dopo il suo rientro in Europa, precisň che gli uccelli del paradiso avevano zampe lunghe circa una spanna, ma non fu creduto.
Sia Gessner che Aldrovandi, viste le aspre polemiche che il vil denaro stava sollevando a mo' di tsunami – pausa: due amici, uno moralista (a) e l'altro no (b): (a) Il denaro č la merda del diavolo!!! (b) O Diavolo, vieni a cagare in casa mia!!! – sia Gessner che Aldrovandi, dicevamo, non possono tacere le accuse nei confronti dell'eresiarca Pigafetta, la prima voce blasfema e competente che cercň di ridimensionare la manucodiata. Gessner cita tutto ciň che il polacco Melchior Guilandinus - alias Melchior Wieland (1519/1520–1589) - aveva scritto nella lunga relazione inviatagli a Padova, senza ovviamente omettere Pigafetta: "[...] praeter unum Antonium Pigafetam, rostro prolixo, & pedibus palmi unius longitudine donari, falsissime affirmantem, quum hoc me hercule longe ab omni veritate sit alienum [...]" (Historia animalium III, 1555, pag. 613)
Aldrovandi, dopo aver descritto le caratteristiche somatiche della manucodiata su cui tutti erano d'accordo – e che facevano cosě passare Aristotele per bugiardo, secondo il quale nessun uccello era senza zampe - chiude il paragrafo con queste parole: "Atque istaec sunt, quae omnibus simul conveniunt: quibus neotericorum peritissimi quique praeter unum Antonium Pigaphettam, qui pedes illis palmum unum longos falsissime tribuit, calculum uno ore adijciunt." (Ornithologia I, 1599, pag. 807)
E le spoglie della manucodiata successivamente portate in Europa, soprattutto dai mercanti olandesi, sembravano contraddire in modo inequivocabile l'affermazione dell'eresiarca. Sorsero cosě le piů curiose leggende su questi volatili. Si credette che gli Uccelli del Paradiso, come vennero chiamati intorno al 1590 dall’olandese Jan van Linschoten, volassero e fluttuassero perennemente ad altissime quote nutrendosi di rugiada e di nettare e che mai si posassero a terra se non per morire. Pertanto non avevano bisogno di piedi. La coppia avrebbe addirittura covato insieme nell’aria, con l’uovo o le uova adagiate in un’infossatura del dorso del maschio e protette dall’addome della femmina.
A dire il vero, la maternitŕ del termine Uccelli del Paradiso universalmente attribuita a Jan van Linschoten - che l’avrebbe partorito intorno al 1590 - potrebbe essere messa in seria discussione, visto che Gessner giŕ nel 1555 chiamava la manucodiata Paradisi avis vel Paradisea. Questi e altri sono i poteri della mancata verifica storica! Ammesso che in casi come questo ne valga la pena.
Poiché le suddette testimonianze esercitavano una sorta di magica seduzione, vi si prestava fede volentieri, benché eminenti studiosi - come Ulisse Aldrovandi - si affannassero a smentirle. Solo nel 1824 uno speziale di bordo, il francese René Lesson, riuscě a scoprire in Nuova Guinea i primi uccelli del paradiso vivi. Si vide allora che non si trattava affatto di creature eteree prive di piedi e viventi nell’aria a spese di nettare e rugiada, bensě di uccelli veri, dotati di zampe robuste.
Alla
scoperta e ai primi entusiasmi per la bellezza di questi uccelli - ora non
piů apodi - seguirono ben presto una sfrenata persecuzione da parte di
cacciatori specializzati e un fiorente commercio con le case di moda
parigine, londinesi e di altre cittŕ. La moda aveva infatti scoperto che le
penne degli uccelli del paradiso costituivano un grazioso ornamento per i
cappellini delle signore. Da secoli, e forse addirittura da millenni, i
Papua![]() in occasione delle loro feste si ornano con le penne di alcune
specie, e se ne servono persino come valuta pregiata per l’acquisto di
mogli e merci. Per fortuna questa caccia tradizionale non era mai riuscita a
turbare l’equilibrio biologico delle diverse specie. Lo sfruttamento del
patrimonio faunistico mediante la caccia organizzata degli Europei,
paragonabile a una febbre dell’oro, e gli atti criminosi condotti
parallelamente a danno degli indigeni, si spinsero a tal punto che nel 1924, quando i prezzi erano comunque precipitati, fu severamente
proibito in Nuova Guinea catturare uccelli del paradiso e possederli. Da
allora solo le istituzioni scientifiche sono
autorizzate alla cattura.
in occasione delle loro feste si ornano con le penne di alcune
specie, e se ne servono persino come valuta pregiata per l’acquisto di
mogli e merci. Per fortuna questa caccia tradizionale non era mai riuscita a
turbare l’equilibrio biologico delle diverse specie. Lo sfruttamento del
patrimonio faunistico mediante la caccia organizzata degli Europei,
paragonabile a una febbre dell’oro, e gli atti criminosi condotti
parallelamente a danno degli indigeni, si spinsero a tal punto che nel 1924, quando i prezzi erano comunque precipitati, fu severamente
proibito in Nuova Guinea catturare uccelli del paradiso e possederli. Da
allora solo le istituzioni scientifiche sono
autorizzate alla cattura.
Prima
di analizzare il testo del 1600 di Aldrovandi - che
giŕ allora non credeva assolutamente all’apodia – e la poesia di
Annibale Bonagente che invece ci credeva, vediamo poche ed essenziali
notizie ornitologiche sull’uccello del paradiso, denominazione attribuita
a gran parte degli Uccelli Passeriformi della famiglia Paradiseidi. Come
giŕ abbiamo detto, la Paradisea apoda, la specie piů nota, deve la
singolare denominazione al fatto che i primi esemplari giungevano in Europa
grossolanamente alterati dalla sommaria imbalsamazione cui venivano
sottoposti dagli indigeni delle isole Aru, comportante, tra l'altro,
l'amputazione delle zampe. I maschi, lunghi poco meno di 50 cm, hanno un
robusto becco diritto di colore grigio-azzurro; zampe anch'esse robuste di
colore rossiccio; due grandi ciuffi di penne estremamente lunghe
giallo-arancio situati ai lati del tronco e le due timoniere centrali
lunghissime e filiformi. Il tronco č rosso-marrone, mentre giallo, verde e
nero compaiono in altre regioni del corpo. Le femmine, prive dei due grandi
ciuffi, sono uniformemente colorate in rosso-marrone. Il maschio, durante il
periodo degli amori, effettua spettacolari parate in cui dispiega tutta la
bellezza del suo piumaggio. Riportiamo solo il nome scientifico di altri
uccelli del paradiso: Paradisea rubra, Paradisea rudolphi, Seleucides
ignotus, Parotia sexpennis, Cinnurus regius, Diphyllodes
respublica, Epimachus fastosus, Lophorina superba e Pteridophora
alberti.

Maschio di uccello del paradiso - Paradisea apoda
dal
Dictionnaire D'Histoire Naturelle di Charles Orbigny - 1849
Il
1°
gennaio 1564 il cardinale Scipione Gonzaga
(1542-1593),
grande amico di Torquato Tasso, fondava a Padova l’Accademia degli
Eterei. Chiamň lo stesso Tasso a farne parte, nonché i migliori
ingegni dello Studio padovano. Per l’impresa dell’Accademia fu dapprima
proposta una figura che viveva perennemente nell’etere, una curiosa
raritŕ ornitologica - l’Avicula Dei, la
manucodiata - che Annibale Bonagente cosě descriveva agli
Eterei:
Miracol
novo a chi sua fama intende:
dove l'aer seren non turba pioggia
nasce un augel che 'n disusata foggia
di puro spirto nutrimento prende.
E
perché vivo in terra unqua non scende,
manca de' piedi, a cui 'l mortal s'appoggia;
e perch'a volo di continuo poggia,
le penne in giro a sostenersi tende.
Tal
di natura aventuroso ardire
schiera di gir al ciel con l'opre accinta
tolga in esempio a l'onorata impresa.
Sperando,
scorta al fin d'alto desire,
ch'ogni caduca voglia a terra spinta
s'erga a l'eterno ben l'anima intesa.
Ma
Aldrovandi, come abbiamo anticipato, non credeva agli uccelli apodi. Infatti
cosě si esprimeva a proposito della raffigurazione di un uccello apode
giuntagli da Pisa su ordine del Granduca di Toscana Ferdinando I
(1549-1609). Il brano č tratto da Ornithologiae
tomus alter (Nicoao Tebaldino, Bologna, 1646)
pagina 629. Il testo latino viene
riportato in calce allo scopo di poter collocare a lato della traduzione l’immagine
inviata ad Aldrovandi.
|
Il
merlo apode indiano |
|
|
|
F.
Malocchio, sovrintendente dell’orto botanico di Pisa, mi ha fatto
pervenire pochissimo tempo fa l’immagine di questo piccolo uccello
su ordine del serenissimo Granduca di Toscana Ferdinando I dei
Medici. Č apode, cioč, č completamente privo di zampe, e ciň per
un fatto naturale, come egli afferma, per cui, a causa della
somiglianza del resto del corpo che esso condivide coi merli,
soprattutto con quello immediatamente precedente, non sarŕ fuori
luogo chiamarlo merlo apode. Dubito parecchio che questo uccello,
come egli dice, sia privo di zampe per cause naturali: dal momento
che il comune maestro Aristotele |
De Merula apode Indica - Cap. XVII - Iconem huiusce aviculae R. P. F. Malocchius horti Pisani praefectus iussu Serenissimi Ferdinandi Medices Magni Hetruriae Ducis nuperrime mihi transmisit. Apos est, id est, pedibus prorsus caret, idque naturaliter, ut ille asseverat, quare ob reliqui corporis similitudinem, quam habet cum merulis, maxime praecedenti proxime, Merulam apodem appellare non erit inconveniens. Num vero naturaliter, quod ille ait, pedibus careat haec avis, non parum addubito: siquidem communis praeceptor Aristoteles animalia cuncta sanguinea non minus, quam quatuor notis moveri attestatur, quare etiam aves animalia nimirum talia totidem notis moveri statuendum erit, alis nempe binis, et totidem pedibus, aut quopiam alio quod horum vicem gerat, ut in manucodiatis cernere est, quibus sagacissima rerum parens natura, quae nunquam exuperat, nunquam deficit, nervos pro pedibus dedit, quibus arboribus, cum opus est, adhaereant. Unde plane affirmaverim, aut pedes avi fuisse ablatos, aut prorsus monstrosam ipsam esse: est autem, quantum ex pictura conijcere queo dodrantem magna, lata digitos tres binis distincta coloribus, capite, ventre, dorso coloris coccinei intensi, et vegeti: alis et cauda nigris (quare Merula porphyromelanos dici posset, a praedominio scilicet, nam pectus, et primum dorsum maculis lunatis virescentibus variant) rostro Merulae, in extremitate recurvo, nigro, infra tamen, qua capiti coniungitur candicante. In oculis nihil apparebat, utpote qui animante exanimata connivebant.
Bufala su bufala
Il merlo apode toscano, raffigurato e descritto grazie ad Aldrovandi, se non apparteneva a una specie attualmente estinta, č verosimilmente una bufala che va a rimpinguare la bufala primaria, cioč la manucodiata. Due bufale alle quale Aldrovandi non credeva assolutamente. Dopo mesi di attente ricerche e di sapiente meditazione, ecco cosa conclude l'Avvocato Giovanni Canali di Parma, esperto ornitologo, che dŕ ragione a Ulisse e che ringrazio di cuore in quanto per me sarebbe stato impossibile giungere alle sue conclusioni:
"Ho valutato disegno e testo. Ho considerato varie specie di paradisee senza trovare somiglianze perfette. Ho altresě considerato altre specie, di colore analogo a quello descritto, ma risultano diverse per alcuni particolari e la morfologia (ad esempio Piranga scarlatta, Piranga olivacea, e Pitoui variabile, Pitohui kirhocephalus). La struttura del soggetto raffigurato ricorda quella di una paradisea. La mancanza di arti inferiori mi fa pensare alla ben nota leggenda del volo perpetuo degli uccelli del paradiso. Posso senz'altro sbagliarmi, ma mi sento di ritenerlo un disegno di fantasia, legato alla leggenda di cui sopra. Curioso di considerare altre ipotesi, cordialmente saluto. - Giovanni Canali (e-mail di domenica 26 agosto 2007)
Piranga scarlatta - Piranga olivacea

sopra: maschio in abito nuziale - sotto: maschio in muta d'eclissi


femmina di Piranga olivacea
Copertina
del n° 250 di Science del 30 ottobre 1992
In alto e al centro un Pitohui kirhocephalus –
Pitoui variabile in italiano
rispettivamente mimetizzante e non mimetizzante
In basso il Pitohui dichrous al quale si č
ispirato il primo Pitohui kirhocephalus per travestirsi
Several species of pitohui, songbirds endemic to New Guinea, contain the potent neurotoxin homobatrachotoxin, apparently as a chemical defense against predators. Chemical defenses have been known to be used by many organisms but not by birds. See page 799. As a possible antipredator adaptation, certain races (top) of the variable pitohui (Pitohui kirhocephalus) mimic the plumage of the hooded pitohui (Pitohui dichrous; bottom), which contains the highest concentration of toxin. A nonmimetic subspecies of the variable pitohui is in the center. [Illustration: John C. Anderton]

Coppia di Paradisea apoda - Daniel Giraud Elliot (1835-1915)
Chiudiamo questo lungo excursus suscitato sě dalla curiositŕ nei confronti della manucodiata, ma soprattutto dalla singolaritŕ della fantasia umana, capace persino di trasformarsi in agente mutageno e di creare il gene dell’apodia.
E l'excursus lo chiudiamo con una carrellata di immagini. Apriamo la rassegna con quelle di Aldrovandi che aveva a disposizione solo cinque soggetti, di cui uno identico a quello di Gessner, che Aldrovandi colloca per ultimo e che battezza come manucodiata vulgaris. A Ulisse mancava un sesto elemento, l'elemento chiave che forse poteva salvare Pigafetta. Ma questa manucodiata - pietra dello scandalo e redentrice dell'eresiarca - forse non aveva ancora messo i piedi in Europa e solo nel 1669 sarebbe stata descritta e raffigurata da Georg Horst nel suo Gesneri redivivi aucti et emendati tomus II.
Vetrina di Aldrovandi


Vetrina di Horst
Si omette la manucodiata di Horst con le fattezze di quella di Gessner -
corrispondente alla manucodiata vulgaris di Aldrovandi – e che
Horst mette in posizione 5.
Chiudiamo invece la sua vetrina con la redentrice di Pigafetta che č in
postazione 6, alla
quale affianchiamo un'immagine quasi identica tratta da Exoticorum libri
decem (1605) di Charles de l'Écluse![]() – alias Clusius – che nella sua lunga e minuziosa descrizione
dell'uccello non cita assolutamente la presenza di zampe.
– alias Clusius – che nella sua lunga e minuziosa descrizione
dell'uccello non cita assolutamente la presenza di zampe.

|
|
|
Manucodiata di Horst nel castello di Gottorff |
Manucodiata apode di Clusius |
Debbo ringraziare Pascal Gratz ![]() per aver trascritto e tradotto la parte essenziale del testo di Georg Horst
relativo al Rex Avium Paradysearum dotato di zampe inaspettatamente
tridattile e contenuto a pagina 368 del suo Gesneri
redivivi aucti et emendati tomus II (1669):
per aver trascritto e tradotto la parte essenziale del testo di Georg Horst
relativo al Rex Avium Paradysearum dotato di zampe inaspettatamente
tridattile e contenuto a pagina 368 del suo Gesneri
redivivi aucti et emendati tomus II (1669):
Seine Füsse sind gestaltet/ und fast so gross als der Krammets oder andern Vögel Füsse: Dann ob zwar meinte Authores, Aldrovandus auch selbsten/ dafür halten/ dass die Paradiesvögel insgemein kein Füss haben/ so ward doch heutiges Tags ein anders durch den Augenschein bewiesen. Dass aber viel dieser Vögel ohne Füsse gesehen werden/ gibt Thuanus die Ursach/ wann er sagt/ dass etliche/ so solche Vögel zu fangen pflegen/ ihnen nicht allein die Füsse/ sondern auch einen guten Theil des Leibes hinweg schneiden/ ... /damit sie dieselbigen als ein grosses Wunderwerck/ nachmals desto theurer verkauffen/ und man sie auch umso viel zierlicher und fuglicher auff den hüten tragen könne/ ...
che in inglese suona cosě:
Its legs resemble those of the Krammet (a juniper thrush) or other birds and are also nearly as big as these: Even if Aldrovandus says that paradise birds don't have legs, it is known today that this isn't true. Jacques-August de Thou gives the answer why many of these birds are seen without their legs, when he says, that the hunters remove the legs of the birds and other parts of their body, .. , to make them look more miraculous, so they achieve a higher price and look better when worn on hats,...
e cosě prosegue Pascal:
Further Horst adds, that the picture was taken after an exemplar stored in Gottorff that still had it's legs. Thus it isn't clear if Horst used Clusius' drawing as a template even though it bears strong resemblance to it. I find this chapter rather interesting, as it shows that Horst researched well into a specific topic that doesn't appear in Gessner's historia animalium.
Quindi,
grazie a Horst e a Pascal, veniamo a sapere che a cavallo tra il 1500 e il 1600 viveva in Francia un altro eretico: Jacques-August de Thou![]() ,
latinizzato in Thusius. Costui era un pigafettiano dall'occhio lungo
capace di recepire la realtŕ venale delle cose pur essendo solo umanista e
giurista – oddio! nominato ministro delle Finanze da Maria de' Medici
madre di Luigi XIII - non
un illustre naturalista neoterico.
,
latinizzato in Thusius. Costui era un pigafettiano dall'occhio lungo
capace di recepire la realtŕ venale delle cose pur essendo solo umanista e
giurista – oddio! nominato ministro delle Finanze da Maria de' Medici
madre di Luigi XIII - non
un illustre naturalista neoterico.
Poi, nel 1669, alla risma degli eretici va ad aggiungersi Georg Horst che osa pubblicare la riproduzione di una manucodiata tutta zampettante conservata nella Kunstkammer – nella stanza delle opere d'arte - del castello di Gottorff, nello Schleswig-Holstein, oggi stato federato della Germania con capitale Kiel e che confina a nord con la Danimarca. Il castello č vicinissimo a Flensburg che č a un tiro di schioppo dalla frontiera del Kongeriget Danmark. Per i pignoli, qualora volessero visitarlo, ecco le coordinate del castello: 54,5116° N - 9,5415° E. I superpignoli saranno lieti di sapere che il castello dista in linea d'aria circa 742 km da Uppsala, in Svezia, dove Linneo morě nel 1778.
7. Zampe
7.1. Polidattilia
|
Po
- polydactyly |
Autosomico
incompletamente dominante
Gruppo di associazione IV - cromosoma 2
Abitualmente
il pollo č dotato di 4 dita, perň
alcune razze ne hanno 5:
Moroseta, Dorking, Faverolles, Houdan e Sultano. Il mancato
intervento dell'apoptosi![]() - ovviamente su base genetica - potrebbe essere in grado di spiegare la morfogenesi di questo fenotipo. La diffusione di questa
caratteristica razziale č dovuta in parte al fatto che gli amatori vanno
matti per ogni stranezza, ma forse dipende anche dal perdurare di una credenza
giŕ diffusa ai tempi di Varrone e Columella, secondo i quali le galline con 5 dita depongono piů uova. Ecco il passo di Columella
tratto dal De re rustica (VIII,2,7-8):
- ovviamente su base genetica - potrebbe essere in grado di spiegare la morfogenesi di questo fenotipo. La diffusione di questa
caratteristica razziale č dovuta in parte al fatto che gli amatori vanno
matti per ogni stranezza, ma forse dipende anche dal perdurare di una credenza
giŕ diffusa ai tempi di Varrone e Columella, secondo i quali le galline con 5 dita depongono piů uova. Ecco il passo di Columella
tratto dal De re rustica (VIII,2,7-8):
|
Eae [aves] sint rubicundae vel infuscae plumae,
nigrisque pinnis: ac si fieri poterit, omnes huius, et ab hoc proximi
coloris eligantur[...] Sint ergo matrices robii coloris, quadratae,
pectorosae, magnis capitibus, rectis rutilisque cristulis, albis
auribus, et sub hac specie quam amplissimae, nec paribus ungulis:
generosissimaeque creduntur, quae quinos habent digitos, sed ita ne
cruribus emineant transversa calcaria. |
Questi
volatili debbono avere piume rosse o nerastre, e le penne nere, e se
sarŕ possibile vengano scelti tutti di questo colore o di un colore
molto simile [...]
Le riproduttrici siano dunque di colore rossiccio, tarchiate,
posseggano un petto largo, la testa grande, la piccola cresta dritta e
rosso splendente, gli orecchioni bianchi, e sotto questo aspetto li
abbiano quanto piů grandi possibile, e non debbono avere le dita
pari: e precisamente sono ritenute molto fertili quelle con cinque
dita, ma non debbono avere speroni che sporgano di traverso sulle
gambe. |
Ma gli allevatori non vanno matti unicamente per le
stranezze anatomiche. Infatti la prelibatezza della Dorking, della Houdan e
della Faverolles puň essere uno dei motivi principali per cui essi si sono
dedicati e si dedicano tuttora con passione alla selezione di queste razze
pentadattile: in questo modo riescono a mantenere senza difetti anche le
caratteristiche anatomiche delle zampe secondo quanto preteso dallo standard
di ciascuna razza.
Ai tempi di Aldrovandi - quindi nel XVI
secolo - la
pentadattilia doveva essere estremamente rara, come egli afferma a pagina 197
del capitolo I,
sottocapitolo Forma et descriptio, del II volume
della sua Ornithologia, talmente introvabile da ribadirlo a pagina
198:
|
Et
Columella etiam Gallinas probat, quae quinque digitos habent, ita
tamen ne cruribus emineant transversa calcaria. Quare quid de eiusmodi
digitis dicam, plane ignoro: cum alioqui pedes pentadactili
[pentadactyli] neque in
avium hoc genere, nec in alio observari, nisi in monstris ex
abundantia materiae videamus: qualis ille pes penctadatilos [pentadactylus] est, quem
mihi olim nescio a quo donatum in musaeo meo reservo. |
Anche
Columella apprezza quelle galline che hanno cinque dita ma che
tuttavia non debbono avere speroni che sporgono di traverso sulle
gambe. Per cui non so assolutamente cosa dire di tali dita, dal
momento che, peraltro, possiamo constatare da un’abbondanza di dati
che zampe pentadattile non si osservano né in questo né in altro
genere di uccelli, se non nei mostri, come č quel piede pentadattilo
che un giorno mi fu dato non so da chi e che conservo nel mio museo. |
|
[...]
Generosissimae, inquit Columella, habentur, quae quinos habent digitos,
sed ita, ne cruribus emineant transversa calcaria; quod Plinius etiam
testatur, cuius verba paulo ante citavimus, ubi nostro aevo eiusmodi
Gallinas haud dari diximus: cur vero tales Gallinae, quibus nempe
calcaria transversa eminent, vitentur, Columella rationem hanc
adiungit, quod id virile signum sit, illae vero ad concubitum
contumaces, dedignentur admittere marem, raroque etiam foecundae
[fecundae] sint,
denique cum incubant, calcis aculeis ova perfrigant [perfringant]. |
[...]
Columella
dice che sono ritenute molto fertili quelle galline dotate di cinque
dita ma che non debbono avere speroni che sporgono di traverso sulle
zampe, cosa testimoniata anche da Plinio, le cui parole abbiamo citato
poco prima, quando abbiamo detto che ai nostri tempi non si
riscontrano assolutamente galline siffatte. Sul perché si debbano
evitare tali galline dotate appunto di speroni che sporgono di traverso, Columella
adduce questa motivazione: si tratta di un segno di mascolinitŕ,
restie invero all’accoppiamento sono sdegnose nell’accettare il maschio,
e sono anche raramente feconde, infine, quando covano, rompono le uova
con gli speroni della zampa. |
Come documentazione iconografica di una pentadattilia
fornitaci da Aldrovandi, ma da lui inspiegabilmente scotomizzata, riportiamo
nuovamente la mostruositŕ 4 giŕ descritta e commentata nel II volume - XXI-2.1.f.

Pullus iste tribus pedibus
natus tertium habebat,
velut ex uropygio enatum.
Perň il misconoscimento di una pentadattilia
č stato perpetrato da Aldrovandi non solo nel caso di questo pulcino. Come
abbiamo appena visto, dimentico - o ignaro? - delle regole scientifiche, che
esigono sempre un adeguato probabilismo, egli č categorico nell’affermare
che le zampe pentadattile non sono presenti né nei polli né in qualsiasi
altro uccello. Eccetto che nei mostri. Ma č proprio in un mostro che
Aldrovandi non si accorge delle cinque dita!
Come se non bastasse, poche pagine prima egli
commette una mancanza ben piů grave: i suoi disegnatori, pittori e incisori,
dotati di grande perizia, ma soprattutto di eccezionale obiettivitŕ, gli
fornivano delle immagini che erano fotocopie del reale, ed egli nemanco si
degnava di gratificarli mettendo a frutto la loro maestria. Infatti, come
vedremo nel prossimo paragrafo dedicato al gene della polidattilia duplicata,
i suoi artisti gli avevano spiattellato un gallo e una gallina che avevano sě
i tarsi impiumati, ma da un corredo di piume che non nascondeva assolutamente
la loro pentadattilia. E Aldrovandi, nel descrivere questa coppia pedibus
pennatis, non fa il minimo accenno alla loro pentadattilia, che
oltretutto, sia dal punto di vista iconografico che genetico, corrisponde
perfettamente a una pentadattilia duplicata. Si limita a dire che avevano le
zampe gialle!
Aldrovandi in certi casi si dimostra un validissimo scienziato, in altre circostanze, come appunto la pentadattilia, č in grado di compromettere irrimediabilmente la sua credibilitŕ. Avremo modo di aggredire ulteriormente il nostro Ulisse su questo importante argomento quando affronteremo nel paragrafo X.8. un dilemma storico e genetico: se la pentadattilia si diffuse in Europa grazie ai Celti oppure ai Romani.
Come abbiamo detto, normalmente i polli posseggono quattro dita per
ciascun piede e solo alcune razze ne hanno cinque. Il
5° dito si sviluppa al di sopra
del 1°
dito normale e prende origine dal
tarsometatarso, ha
lunghezza maggiore rispetto al 1° dito sottostante, non deve toccare il suolo
e deve essere rivolto all’insů.
Questa situazione anatomica č quella ideale, prevista dallo standard,
ma di fatto la polidattilia ha un’espressione molto variabile, frustrando
cosě talora non poco gli allevatori.
La pentadattilia da gene Po
come pretesa dall’American Standard of Perfection - 1993
La polidattilia č presente all’atto della schiusa, e in effetti č
giŕ visibile al quinto giorno d’incubazione (Kaufmann-Wolf, 1908);
puň venire soppressa sottoponendo gli embrioni a basse temperature a 14 giorni
d’incubazione (Sturkie, 1943;
Warren, 1944).

Scheletri
di zampa di embrioni di Silky giapponese (A, C, E, G, I) e di Livorno
bianca (B, D, F, H, J) in momenti compresi tra il 29° e il 42° stadio di
sviluppo secondo la codifica di Hamburger
& Hamilton (1951)![]() .
Quattro dita cartilaginee erano visibili sia nell'embrione di Silky che in
quello di Livorno allo stadio 29 (6° giorno d'incubazione). Come indica la
punta della freccia, a partire dallo stadio 30 e negli stadi successivi la
zampa di Silky manifesta un dito supplementare che si biforca dalla radice del
1° dito. – Immagine tratta da Skeletal
analysis and characterization of gene expression related to pattern formation
in developing limbs of Japanese Silkie Fowl
di Kenjiro
Arisawa, Shigenobu Yazawa, Yusuke Atsum, Hiroshi Kagami
and
Tamao Ono (The
Journal of Poultry Science,
43
:
126-134,
2006) - Questi studiosi hanno messo in evidenza un fatto importante. Nell'embrione di Silky, e non in quello di Livorno,
si
č potuto osservare a livello dell'abbozzo dell'ala la comparsa di un dito
soprannumerario cartilagineo - un 4° dito - che
nel
momento compreso tra gli stadi 32 e 36 si biforcava dalla radice del 2° dito,
ma allo stadio 37 e nei successivi stadi questo abbozzo non era piů visibile
(penso si tratti di una scomparsa dovuta verosimilmente ad apoptosi).
Vale la pena rendere disponibile il loro studio in formato PDF
.
Quattro dita cartilaginee erano visibili sia nell'embrione di Silky che in
quello di Livorno allo stadio 29 (6° giorno d'incubazione). Come indica la
punta della freccia, a partire dallo stadio 30 e negli stadi successivi la
zampa di Silky manifesta un dito supplementare che si biforca dalla radice del
1° dito. – Immagine tratta da Skeletal
analysis and characterization of gene expression related to pattern formation
in developing limbs of Japanese Silkie Fowl
di Kenjiro
Arisawa, Shigenobu Yazawa, Yusuke Atsum, Hiroshi Kagami
and
Tamao Ono (The
Journal of Poultry Science,
43
:
126-134,
2006) - Questi studiosi hanno messo in evidenza un fatto importante. Nell'embrione di Silky, e non in quello di Livorno,
si
č potuto osservare a livello dell'abbozzo dell'ala la comparsa di un dito
soprannumerario cartilagineo - un 4° dito - che
nel
momento compreso tra gli stadi 32 e 36 si biforcava dalla radice del 2° dito,
ma allo stadio 37 e nei successivi stadi questo abbozzo non era piů visibile
(penso si tratti di una scomparsa dovuta verosimilmente ad apoptosi).
Vale la pena rendere disponibile il loro studio in formato PDF![]() .
.
Warren (1944) ha
proposto tre distinti modi di manifestarsi di questo tratto. Talora ciascuna
di queste modalitŕ espressive non si realizza perfettamente, dando luogo cosě
a una vasta gamma di espressioni anatomiche. Il primo fenotipo indicato da
Warren consiste nell’aggiunta di un lungo dito al di sopra del primo dito
normale; la seconda espressione č rappresentata dalla fissurazione del dito
soprannumerario; con frequenza minore si verifica infine la perdita del primo
dito normale. Con questa ipotesi di lavoro egli fu in grado di dare una
spiegazione razionale a quattro dei fenotipi da lui osservati.
Un quinto fenotipo, che Warren (1944) chiamň
polifalangia,
consisteva in un primo dito dotato di una falange addizionale: sebbene questi
soggetti avessero solamente quattro dita come č la norma, la polifalangia
compariva solamente nel gruppo geneticamente polidattilo, e cosě sembrň
trattarsi di una polidattilia modificata. Warren (1944)
studiň ulteriormente la condizione
di polifalangia in ceppi polidattili puri e concluse che essa si comportava
come se venisse controllata da un gene autosomico dominante insieme ad alcuni
fattori ignoti causa di variabilitŕ della sua dominanza.
La polidattilia monolaterale, nota anche come eterodattilia,
č stata osservata da molti ricercatori (Bond, 1920,
1926; Punnett e Pease, 1929;
Warren, 1944):
il numero di dita per ciascun piede era diverso, con il piede sinistro
abitualmente fornito di dita soprannumerarie e il piede destro dotato delle
normali quattro dita; ma ambedue i piedi potevano essere polidattili, perň
con fenotipo diverso. Con una selezione condotta per piů di sette anni
Landauer (1948) fu
in grado di elevare dal 13 al 33%
la proporzione dei soggetti
eterodattili a sinistra, ma a spese di quelli con polifalangia che quasi
scomparvero dal suo ceppo.
Molti studiosi (Bond, 1926;
Punnett e Pease, 1929;
Landauer, 1948;
Warren, 1944)
hanno fornito ampie informazioni sull'ereditarietŕ della polidattilia:
sebbene sia un tratto
complesso, influenzato
da geni modificatori e soppressori,
la sua ereditarietŕ di base č quella di un gene autosomico incompletamente
dominante. Dunn e Jull (1927) hanno
assegnato a questo locus il simbolo Po.
Esiste tuttavia un inconveniente nell'ereditarietŕ di questo tratto: pare
che alcune razze polidattile, cosě come alcune razze non polidattile, siano
dotate di geni soppressori che possono inibire completamente l'espressione
della polidattilia in soggetti genotipicamente polidattili
(Punnett and Pease, 1929;
Hutchinson, 1931;
Warren, 1944).
Affinché gli allevatori, di fronte a queste incognite di carattere
genetico, non vengano colti da scoraggiamento qualora avessero deciso di
allevare razze pentadattile, cito la risposta di Jean-Claude
Périquet, da me espressamente interpellato su eventuali difficoltŕ
d’allevamento di soggetti pentadattili:
“Non esistono difficoltŕ particolari nell’ottenere con una buona
selezione delle belle dita nelle razze a 5 dita. Certamente, in queste razze
si riscontrano delle unghie doppie, un primo e un quinto dito troppo
ravvicinati, talora degli accenni a un sesto dito, un’unghia mancante: ma
questi soggetti debbono essere eliminati dalla selezione.”
(E-mail del 26-6-2003:
“Il n'y a pas de difficultés particuličres pour
obtenir de bons doigts dans les races ŕ 5 doigts
par une bonne sélection. Certes, on trouve dans ces races des ongles doubles,
des 4e et 5e doigts trop serrés, parfois des débuts de 6čme doigt, un ongle
manquant: mais ces sujets doivent ętre éliminés de la sélection.”
E-mail del 18-8-2003:
“Ce
que nous nommons 4čme et 5čme doigts, ce sont les 2 doigts ŕ l'arričre qui
ne touchent pas le sol.”)
Pentadattilia
Apelle Dei - 16 gennaio 1890
L’aumento numerico delle dita rispetto alla norma č
detto polidattilia, e quando un individuo con solo 2 dita, come lo struzzo,
giunge ad avere 5 dita, la sua polidattilia č ovviamente detta pentadattilia.
La collocazione della polidattilia in seno alle mostruositŕ non č da tutti
accettata, in quanto neppure per l’uomo costituisce una deformitŕ marcata,
e siccome non influenza in modo apprezzabile il funzionamento degli arti, non
dovrebbe trattarsi di una mostruositŕ, ma di un’anomalia, e quindi di una
semplice variazione rispetto a quanto si osserva abitualmente.
Si
tratterebbe di un
ritorno verso forme ancestrali, anche se esiste un’importante differenza tra l’atavismo
del cavallo, del cane e del pollo nei confronti dell’atavismo dell’uomo.
Nel pollo la polidattilia non va mai oltre un 5° dito. Č stato affermato che
si tratta di una duplicitŕ o sdoppiamento dell’alluce, tuttavia ciň non
parrebbe vero.
Negli
uccelli il numero normale di dita puň essere inferiore a 4, ma non č mai
superiore a 5. In essi č scomparso il 5° dito, o mignolo, per ragioni di
statica. La disposizione normale delle dita di un pollo č data dall’alluce
volto all’indietro e diretto verso l’interno, dotato di 2 falangi e privo
di tendine del lungo estensore. Esistono poi 3 dita volte in avanti, dotate, a
partire dal lato mediale, di 3, 4 e 5 falangi. Queste dita sono mosse dal
tendine del lungo estensore.
|
|
Fig.
X. 8 - Pianta del piede sinistro in gallina pentadattila a,
b, c, d: alluce, II, III, IV dito normali |
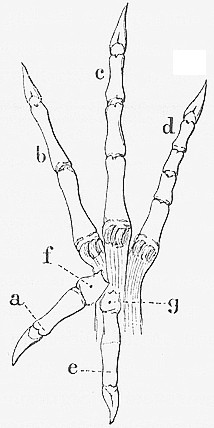
Il
5° dito, scomparso negli uccelli, potrebbe fare la sua ricomparsa nei polli,
e quando ricompare č volto all’indietro, essendo questa la tendenza delle dita poste alla
periferia della zampa. Per cui non ci si deve meravigliare che il mignolo
tenda a ricomparire orientato come l’alluce. Per lo piů il dito
soprannumerario č dotato di 3 falangi (l’alluce ne ha solo 2) e rispetto
all’alluce presenta delle inserzioni tendinee.
Nell’uomo,
e negli animali dotati di 5 dita a livello degli arti toracici e pelvici, il
numero di dita soprannumerarie puň essere maggiore di uno. Nei polli invece
č sempre uguale a uno.
Quanto contenuto nelle precedenti righe č frutto delle
osservazioni su Pollo, Pavone e Faraona da parte di Apelle Dei, il quale, il
16 gennaio 1890, in Siena, comunicň i suoi dati alla Regia Accademia dei
Fisiocritici. Il Dei, da vero scienziato, lasciava aperta la possibilitŕ di
essere contestato, e la contestazione proviene proprio dai geni che
descriveremo tra poco.
7.2. Polidattilia duplicata
|
Pod - duplicate
polydactyly |
Autosomico
dominante, allele di Po
Gruppo di associazione IV - cromosoma 2
Questo tipo di polidattilia č causa di una maggiore
duplicazione del 1° dito
e presenta un’espressivitŕ piů ampia ![]() .
.
Warren (1941) riferě
di un tipo nuovo di polidattilia che con test di allevamento dimostrň essere
autosomica dominante e allele di Po. Questo allelismo fu
successivamente comprovato, sempre dallo stesso Warren (1944), usando
test di linkage con i loci dello sperone duplice e degli speroni multipli.
Questo tipo di polidattilia, che Warren (1941)
chiamň duplicata, causava
molto piů frequentemente una duplicazione del primo dito, con una piů ampia
variazione di espressivitŕ.
Le espressioni piů frequenti erano:
- una condizione con 6 dita
nella quale erano presenti un 1°
dito normale e una fissurazione del
dito addizionale
- una condizione con 5 dita
nella quale il 1° dito
normale era sostituito da un dito lungo con vari gradi di fissurazione; quindi
non si trattava di polidattilia da dito soprannumerario, ma di una
fissurazione di grado variabile a carico di un primo dito abnormemente lungo.
In alcuni casi estremi erano presenti dita soprannumerarie lungo la
zampa fino all’altezza dell’articolazione del ginocchio, mentre in altri
casi era presente una quasi completa duplicazione del piede e del
tarsometatarso. I casi estremi di solito non sopravvissero.
Fu frequentemente osservata un’asimmetria di espressione tra i due
piedi. Una duplicazione del secondo dito della mano era evidente nei soggetti
con polidattilia duplicata, ma non nei soggetti polidattili ordinari.
Una documentazione iconografica di questo tipo di polidattilia ci viene involontariamente offerta da Ulisse Aldrovandi, che nel II volume della sua Ornitholgia descrive il gallo e la gallina pedibus pennatis senza perň metterne in evidenza la palese condizione a cinque dita, frustrando per una seconda volta - la prima fu quella del pulcino mostruoso e pentadattilo - frustrando, dicevamo, la preziosa attivitŕ del suo stuolo di disegnatori, pittori e incisori, dotati di grande perizia, ma soprattutto di eccezionale obiettivitŕ fotografica.
Pullus iste tribus pedibus
natus tertium habebat,
velut ex uropygio enatum.
Le immagini in bianco e nero dell’Ornithologia
relative a questa coppia di polli sono disponibili - con qualche piccola
variazione - anche in versione acquarello ![]() -
- ![]() che nel novembre 2004 sono
state autenticate grazie alla Dottoressa Rita Giordano (Museo Aldrovandi -
Bologna). Esse
non necessitano di speciali didascalie, essendo i particolari piů che
evidenti.
che nel novembre 2004 sono
state autenticate grazie alla Dottoressa Rita Giordano (Museo Aldrovandi -
Bologna). Esse
non necessitano di speciali didascalie, essendo i particolari piů che
evidenti.
Le rappresentazioni a colori sono
catalogate come Gallus et Gallina hirsutis pedibus.
Il merito della scoperta di questo
imperdonabile svarione di Aldrovandi č tutto della Dottoressa Irina
Moiseyeva di
Mosca la quale si č accorta della situazione pentadattila esaminando le
immagini a colori - quelle ad acquarello - contenute in Poultry of the World (1996) di Loyl
Stromberg. La scoperta di Irina mi veniva comunicata con una e-mail del
2-11-2004 con queste parole:
«Please, look at the book by
Stromberg "Poultry of the World", p. 68-69. Are these birds with
five toes? Are they from Aldrovandi's collection of drawings? How these
drawings are in accordance with the text in our paper?»
A queste allarmate e allarmanti parole
di Irina ha fatto seguito tutta la bagarre con Bologna nonché lo
scoppio della bomba sull’inaffidabilitŕ di Aldrovandi per ricerche di
genetica storica.
Fu cosě che il gallo e la gallina hirsutis pedibus assursero al ruolo di pietra dello scandalo.

Ed ecco il testo di Aldrovandi di pagina 313 che descrive la pietra
dello scandalo, senza accennare assolutamente al quinto dito, ma solo al
colore giallo delle zampe. Leggere per credere!
|
de
gallo, et gallina |
Capitolo V il
gallo e la gallina dalle zampe impiumate |
|
Sunt
quoque in hoc avium genere pedibus quandoque hirsutis, quales sunt,
quos nunc damus, Gallus, et Gallina, Gallus collo, et dorso erat
castanei coloris. Alae primum nigrae albicantibus maculis decoratae,
dein nigrae erant, remigae candidae extra, intus nigrae. Mentum,
pectus, venter coxae, tibiae nigra, albicantibus maculis insignita.
Pedes lutei, crista duplex non admodum magna rostrum luteum: paleae
magnae; cauda partim albis, partim nigris pennis constabat. Gallina
tota flavescebat, feri [fere] atris ubique, si solum collum demas,
maculis, modo parvis, modo magnis, at ubique oblongis conspersa;
cristam habebat omnium minimam et paleas admodum breves, rostrum, et
pedes luteos. |
Anche
in questo genere di volatili vi sono dei soggetti che talora hanno le
zampe irsute, come lo sono quelli che adesso mostriamo, un gallo e una
gallina; il gallo aveva il collo e la schiena di colore castano. Le
ali nella parte anteriore erano nere ornate da macchie biancastre,
quindi erano nere, le remiganti erano bianche all’esterno, nere
all’interno. La gola, il petto, il ventre, le cosce e i tarsi erano
neri, fregiati di chiazze biancastre. I piedi erano gialli, la cresta
doppia non eccessivamente grande, il becco giallo: i bargigli grandi,
la coda era costituita da penne in parte bianche e in parte nere. La
gallina era tutta quanta fulva, cosparsa dappertutto, se si esclude
solamente il collo, da macchiettature che erano quasi nere, ora
piccole, ora grandi, ma ovunque si presentavano allungate, aveva la
cresta piů piccola di tutte e i bargigli molto corti, il becco e i
piedi gialli. |
7.3. Polidattilia recessiva
|
po-2
- recessive polydactyly |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Un
tipo recessivo di polidattilia fu scoperto in alcune Rhode Island Red nelle
quali il 22% dei soggetti
interessati aveva 5 dita,
il 31% aveva 6
dita, e il rimanere 47%
aveva la mancanza del dito
posteriore che era sostituito da 2-3 dita
addizionali sul tarsometatarso (Ibe e McGibbon, 1980).
La mortalitŕ dei pulcini affetti era circa il doppio di quella dei
soggetti normali. Circa il 25% morě
allo stadio di embrione tardivo, mentre un ulteriore 17%
morě intorno a sei settimane d’etŕ.
I soggetti ancora vivi oltre le sei settimane abitualmente mostrarono delle
deformitŕ alle gambe.
Ibe e McGibbon (1980) proposero
il simbolo por per questo tratto; tuttavia essi non suggerirono trattarsi di un
allele del locus della polidattilia dominante (Po) e cosě a questo
locus č stato successivamente assegnato il simbolo po-2
(Somes, 1981).
7.4. Brachidattilia
|
By - brachydactyly |
Autosomico
incompletamente dominante
Gruppo di associazione sconosciuto
Questo
gene interessa
il 4° dito del
piede, quello piů esterno, che nel pollo normale č piů lungo di circa il
10% rispetto al 2° dito, ed č formato da 5 falangi. In quest’anomalia il
dito piů esterno uguaglia in lunghezza quello interno, o č persino piů
corto. Gli omozigoti sono privi di alcune falangi e dell’unghia
corrispondente, ma non sempre. L’espressione estrema č rappresentata da 2
sole falangi con assenza dell’unghia. Gli eterozigoti presentano anomalie
minori. Pare che siano i maschi a mostrare le anomalie di maggior entitŕ o il
fenotipo estremo.
La
malformazione č giŕ riconoscibile nell’embrione di 9-10 giorni, ed č piů
frequente nelle razze a tarsi impiumati, come Brahma e Cocincina. Warren
(1940) affermava l’esistenza di una relazione inversa fra entitŕ d’impiumamento
e lunghezza relativa del 4° dito: i soggetti dai tarsi meglio impiumati sono anche quelli
che hanno il 4° dito piů corto. A causa di queste correlazioni, Schmalhausen ha proposto
un solo gene come causa dei differenti fenotipi, ipotizzando che la crescita
delle piume ai tarsi possa interferire con un normale sviluppo del dito.
Tuttavia, esiste anche l’ipotesi secondo cui i due tratti si trasmettano in
modo indipendente, per cui non esisterebbe uno stretto linkage col gene dell’impiumamento
dei tarsi, quanto piuttosto un processo comune alle due caratteristiche
fenotipiche che si svolge durante la vita embrionale.
Di nuovo si č fatta strada l’ipotesi secondo cui impiumamento e brachidattilia siano condizionati da un solo identico gene, in quanto l’impiumamento migliore dei tarsi sembra dovuto alla compartecipazione di due differenti loci, e non si puň escludere, anzi pare verosimile, che uno di questi due geni e il gene della brachidattilia siano la stessa cosa. Danforth č giunto addirittura ad affermare che ambedue i geni implicati nell’impiumamento dei tarsi sono in grado di provocare la brachidattilia.

Maschio
eterozigote di Marans nana all'etŕ di 2 mesi - 2009
allevato da Elly Vogelaar![]() a
Voorschoten nella provincia dell'Olanda Meridionale.
a
Voorschoten nella provincia dell'Olanda Meridionale.
Evidente
brachidattilia al IV dito del piede
con impiumamento dei tarsi piů accentuato del solito.
foto di Dirk de Jong – Ijsselstein nella provincia di Utrecht
7.5. Anomalie del dito medio e di tutte le dita
|
Anomalie
del dito medio e di tutte le dita |
Ereditarietŕ
non ancora chiarita
La
confusione riguardo l’esatto genotipo dei tarsi impiumati si accresce, in
quanto l’impiumamento dei tarsi viene classificato secondo fenotipi diversi,
uno dei quali č caratterizzato dalla mancanza
dell’unghia del dito medio, o 3° dito, che si presenta anche nudo, salvo poche piume sul lato
esterno, con rimanente impiumamento del tutto normale. Quest’anomalia
potrebbe rientrare in questo paragrafo e non in quello precedente. Spesso i
pulcini perdono le unghie in quanto circondate da pallottole di fango,
poiché, se queste vengono rimosse dall’allevatore o da un trauma, possono
portare con sé l’unghia inglobata. La mancanza di unghia per questo motivo
puň essere facilmente differenziata da quella dovuta al gene By, in
quanto la lunghezza di tutte le dita, specie di quelle piů esterne, č
normale.
Le
anomalie del dito medio sono frequenti. Tuttavia la loro base genetica non č
ancora stata compresa appieno. Nel 75% dei soggetti portatori di un’anomalia
di questo dito si riscontra un’assenza dell’unghia, oppure l’unghia puň
essere accorciata o variamente deviata in una o in ambedue le zampe. Talora l’unghia
si limita a essere curvata da un lato.
In casi piů severi tutte le dita sono anormali, con
anomalie anche a carico delle ossa. Questa situazione fu battezzata da Hutt
come osteodistrofia
ungueale,
spesso ritenuta ingiustamente accidentale, ragion per cui essa non riesce a
scomparire da un ceppo, in quanto i soggetti che ne sono affetti vengono poi
fatti regolarmente riprodurre. Per cui, occhio ai piedi! specie se nascosti da
abbondanti piume!
7.6. Zampa d’anatra
|
Zampa d’anatra |
Ereditarietŕ non ancora studiata
Č uno dei difetti piů frequenti nel pollo, con alluce
diretto anteriormente, per cui sembra che il soggetto cammini sui tacchi
come gli
anatidi, nei quali tuttavia il primo dito č diretto posteriormente.
La genetica di questo difetto non č stata studiata, ma pare si tratti di un
carattere recessivo.
7.7. Dita torte congenite
|
crt - congenital
crooked toes |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Ha un’ereditarietŕ uguale nei due sessi, con penetranza
incompleta influenzata da geni modificatori variabili da un ceppo all’altro.
Un’anomalia morfologica delle dita, conosciuta come dita torte o arricciate,
non č rara, in cui fattori ambientali, come la deficienza di riboflavina, l’allevamento
su pavimento levigato, l’affollamento sotto le fonti di calore, sono in
grado di causare questa deformitŕ. Esiste tuttavia anche la malformazione su
base genetica, indistinguibile da quella dovuta a puri fattori ambientali.
Zampa
d'oca del 1556
descritta da Licostene
![]()
Ciň che stiamo per affrontare non ha nulla a che vedere dal punto di vista fenotipico con la mutazione zampa d'anatra appena descritta. L'unico addentellato č puramente lessicale. E siccome disquisiremo della membrana interdigitale dei palmipedi, dire zampa d'anatra o zampa d'oca č del tutto equivalente.
Forse stiamo per dedicarci a una mutazione del pollo ormai scomparsa. Probabilmente nel 1992 - per la prima e unica volta - mi č capitato di vedere questo fenotipo in una gallina. Ho interpellato nel 2006 l'allevatore di questa gallina dotata di un'ampia membrana interdigitale ad ambedue le zampe, ma egli purtroppo disconosce il mio reperto visivo, per cui non posso continuare a millantarmi sostenendo di avere una memoria pari a quella di una macchina fotografica. Perň, credetemi, quella gallina me la ricordo.
Licostene - alias Conrad Wolffhart (1518-1561) – nel suo Prodigiorum ac ostentorum chronicon (1557) č stato il primo a riferire la notizia di galline con zampe d'oca:

anno Domini 1556
Augustae Rauracorum inter Basileam ac Rinfeldam oppida, gallinae anserum pedibus inventae sunt.
Ad Augst![]() ,
che si trova tra Basilea e Rheinfelden, si sono trovate delle galline con
zampe d'oca.
,
che si trova tra Basilea e Rheinfelden, si sono trovate delle galline con
zampe d'oca.
Se si fosse trattato di un'unica gallina, Licostene l'avrebbe segnalato, come ha fatto nel caso di un solo pollo con quattro zampe citato insieme a un contadino dotato di 6 dita ad ambedue le mani:

anno Domini 1556
Basileae Rauracorum agricolam in utraque manu sex digitos iustae magnitudinis ac aptae proportionis habentem, & gallinae pullum quatuor pedibus, vidimus.
Ho visto a Basilea un contadino della stirpe dei Rauraci che ad ambedue le mani presentava sei dita di giusta grandezza e adeguatamente proporzionate, nonché un pollo con quattro zampe.
Possiamo quindi presumere che per Augst non si trattasse - come nel mio caso - di una singola gallina, bensě di un certo numero di galline con zampe d'oca, tanto da poter pensare a un'anomalia di tipo genetico trasmissibile ereditariamente.
Nell'appendice
a Ornithologiae tomus tertius ac postremus (1603) Aldrovandi allega quattro nuove mostruositŕ riscontrate in pulcini di gallina![]() e questi quattro soggetti vengono debitamente documentati anche nel suo Monstrorum
historia edito postumo nel 1642 grazie a Bartolomeo Ambrosini
e questi quattro soggetti vengono debitamente documentati anche nel suo Monstrorum
historia edito postumo nel 1642 grazie a Bartolomeo Ambrosini![]() (1588-1657).
(1588-1657).
Perň le
galline anserum pedibus riferite da Licostene, pur rappresentando
un'insolita mostruositŕ, sono inspiegabilmente assenti in ambedue i trattati
di Aldrovandi, nonostante le numerose citazioni di polli e di non polli
desunte da Licostene, sia sotto forma di notizie che di immagini, talora
spavaldamente manipolate![]() .
.

Gallo
con 4 zampe e gallina con zampe d'oca
di Johann-Georg Schenck - 1609
Invece la
gallina dai piedi anserini viene debitamente annoverata da Johann-Georg
Schenck - figlio di Johann
Schenck von Grafenberg![]() - nel suo Monstrorum historia (1609) unitamente ad altre immagini
desunte da Licostene. La gallina di Schenck non č identica nei dettagli a
quella di Licostene, ma si puň tranquillamente affermare che si tratta di
un'immagine speculare.
- nel suo Monstrorum historia (1609) unitamente ad altre immagini
desunte da Licostene. La gallina di Schenck non č identica nei dettagli a
quella di Licostene, ma si puň tranquillamente affermare che si tratta di
un'immagine speculare.
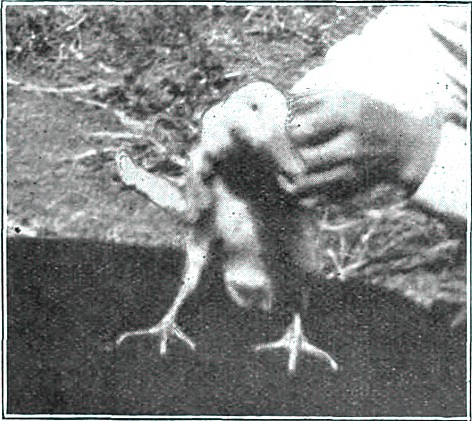
Anatra di Morganville fotografata da John Henry Schaffner - 1921
Nel 1921
il botanico John Henry Schaffner dell'Ohio State University descriveva e
forniva la documentazione fotografica di un'anatra con zampe di pollo![]() nata in seno a uno stuolo di 50 anatre dalle zampe normali allevate da Louis
Weiss nei pressi di Morganville nel Kansas.
nata in seno a uno stuolo di 50 anatre dalle zampe normali allevate da Louis
Weiss nei pressi di Morganville nel Kansas.

Anseriformi con zampe di pollo
Se le immagini di questi anseriformi con zampe di pollo - volutamente dotati da madre natura anche di un becco che ricorda quello del pollo - fossero giunte nelle mani di Licostene, Aldrovandi e Schenck dopo essere stati faticosamente catalogati tra gli anseriformi, credo che questi autori rinascimentali li avrebbero subito schedati tra le mostruositŕ, mentre dal punto di vista biologico non si tratta assolutamente di anseriformi mostruosi anche se inabituali.
Naturalmente nel XVI secolo la possibilitŕ di venire a conoscenza di siffatti uccelli dipendeva dal progressivo ampliarsi delle esplorazioni e dalla diligenza dei cronisti, nonché dall'accessibilitŕ dell'habitat che, nel caso degli Animidi, č piuttosto faticoso da raggiungere. Cosě nel XVI secolo questi anseriformi non erano ancora approdati in Europa.
Puň un pollo presentare zampe d'oca? E di converso: puň un'anatra – tipo quella di Morganville – ritrovarsi con zampe di pollo con cui dover camminare e nuotare?
Teoricamente
e praticamente sě, perň per meccanismi estranei al consueto sviluppo
embrionale, mentre in via del tutto fisiologica presentano zampe di pollo i
tre appartenenti alla famiglia Animidi![]() - Anhima cornuta, Chauna torquata e Chauna chavaria -
nonché l'unico rappresentante della sottofamiglia Anseranatini, la Anseranas
semipalmata o Oca gazza o Oca semipalmata
- Anhima cornuta, Chauna torquata e Chauna chavaria -
nonché l'unico rappresentante della sottofamiglia Anseranatini, la Anseranas
semipalmata o Oca gazza o Oca semipalmata![]() .
.
Nel pollo
esiste la mutazione talpa![]() - letale per l'embrione - che determina mani e piedi simili a quelli di una
talpa, per cui non si puň escludere che per un periodo limitato di tempo nei
polli di Augst sia comparsa la mutazione zampa d'oca, priva perň di azione
letale sull'embrione, e che successivamente sia andata perduta, come accade
quotidianamente per parecchie mutazioni genetiche anche non letali.
- letale per l'embrione - che determina mani e piedi simili a quelli di una
talpa, per cui non si puň escludere che per un periodo limitato di tempo nei
polli di Augst sia comparsa la mutazione zampa d'oca, priva perň di azione
letale sull'embrione, e che successivamente sia andata perduta, come accade
quotidianamente per parecchie mutazioni genetiche anche non letali.
Per
affermare che anche il genere Gallus possa presentare un'estesa
membrana interdigitale bisogna supporre che nell'embrione sia venuto a mancare
il meccanismo biologico che fa scomparire tale membrana, cioč un blocco
locale dell'apoptosi![]() ,
detta anche morte cellulare programmata.
,
detta anche morte cellulare programmata.
Credo invece che negli anseriformi dai piedi di pollo accada l'opposto di un blocco dell'apoptosi: in essi la morte cellulare programmata č fisiologicamente attiva nel determinare l'assenza di membrana interdigitale.
Perň per gli Animidi e per l'Oca gazza forse non č stato possibile effettuare studi embriologici a causa delle poche uova disponibili rispetto all'elevato numero richiesto, studi ai quali si č invece brillantemente prestato il pulcino di gallina, la quale di uova ne depone a iosa. Comunque, č facile arguire che in questi anseriformi sia attiva un'apoptosi fisiologica, cosě come č fisiologica quella di un normale pulcino.
Conosco solo un dato della biologia sperimentale relativo alla sindattilia nel pollo, e anch'esso si basa sull'apoptosi, come affermano Michael Wride & Leon Browder nella loro specifica monografia: "If chick limb mesoderm is combined with duck ectoderm, PCD [programmed cell death] fails and the digits remain joined (Saunders, 1966)." Cioč: se il mesoderma di una zampa del pulcino viene combinato con l'ectoderma dell'anatra, l'apoptosi viene meno e si ha la sindattilia. Per cui alla nascita osserveremo in questo pulcino una zampa d'oca o, se preferiamo, una zampa d'anatra.
Quindi, non essendo assolutamente bizzarre queste ipotesi morfogenetiche circa un pollo con zampa d'oca e un anseriforme con zampa di pollo, esse darebbero credito alla serietŕ storiografica di Licostene.
Per ora
– e spero anche per il futuro – studi sugli arti umani come quelli
condotti nell'embrione di pollo non stanno ancora riempiendo le pagine delle
riviste scientifiche. Accontentiamoci quindi di ipotesi: non possiamo
escludere che la mancata attivazione di una fisiologica apoptosi possa
giustificare l'esadattilia umana![]() ,
cosě come pare possa spiegare la polidattilia del pollo
,
cosě come pare possa spiegare la polidattilia del pollo![]() .
.

Raffaello
- Sposalizio della Vergine - 1504
Esadattilia del piede sinistro di San Giuseppe
magari
suggerita da quella del Perugino
Ed ecco
che l'esadattilia del Rauraco di Basilea riferita da Licostene va a tenere
compagnia a quella del X secolo aC citata da Samuele (II 21,20-22) finita
cadavere per mano di un certo Gionata, nonché a quelle del XV secolo dello Sposalizio
della Vergine del Perugino e di Raffaello recentemente diagnosticate da
Andrea Bertolazzi![]() (2006), e poi a quelle del XVII secolo di San Marco e San Luca raffigurate nei dipinti della
(2006), e poi a quelle del XVII secolo di San Marco e San Luca raffigurate nei dipinti della
|
sommario |
top |
avanti |
|
|
|